

Uracil excision by endogenous SMUG1 glycosylase promotes efficient Ig class switching and impacts on A:T substitutions during somatic mutation
- PMID: 24771041
- PMCID: PMC4158878
- DOI: 10.1002/eji.201444482
Uracil excision by endogenous SMUG1 glycosylase promotes efficient Ig class switching and impacts on A:T substitutions during somatic mutation
Abstract
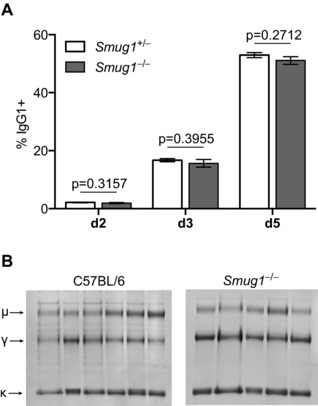
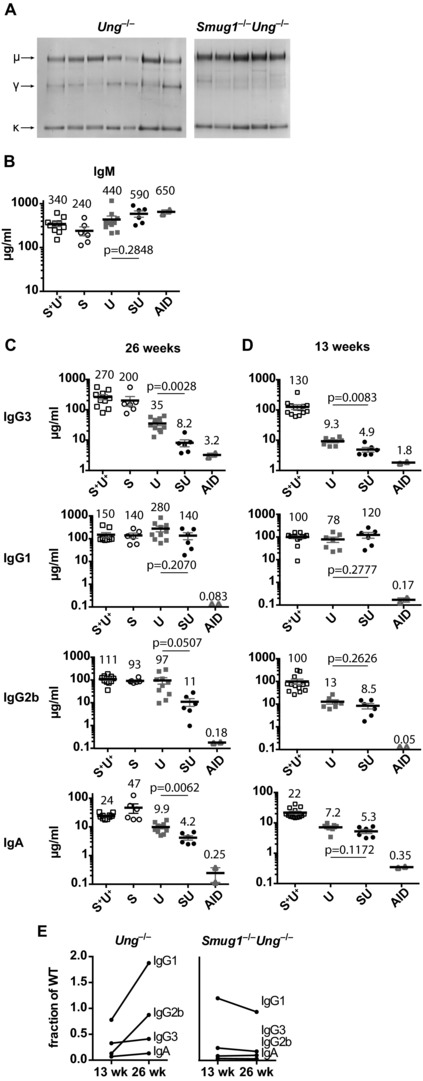
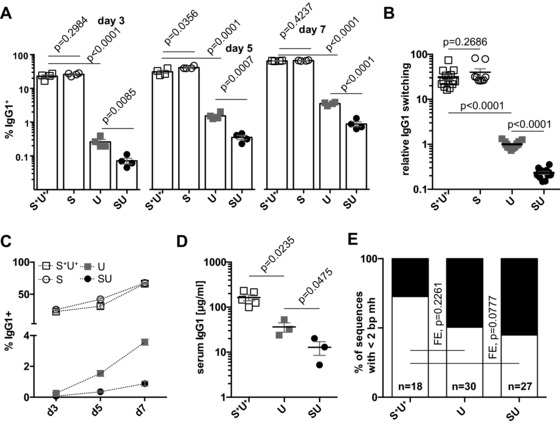
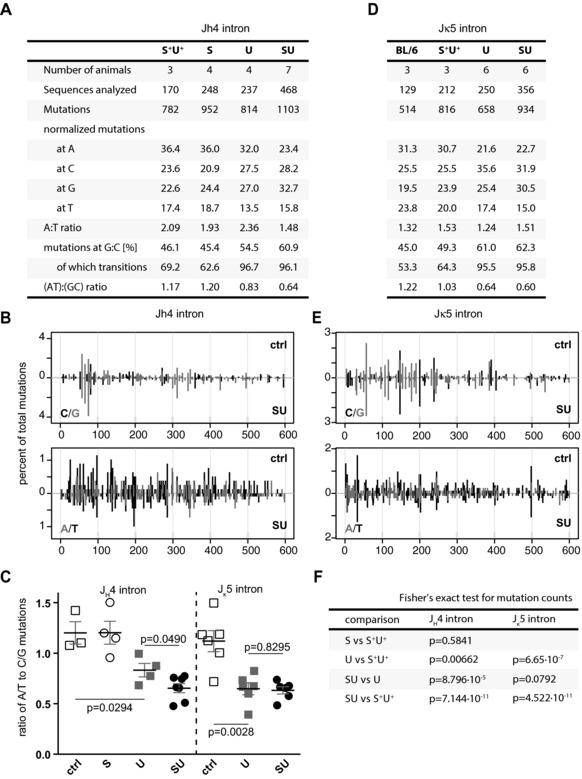
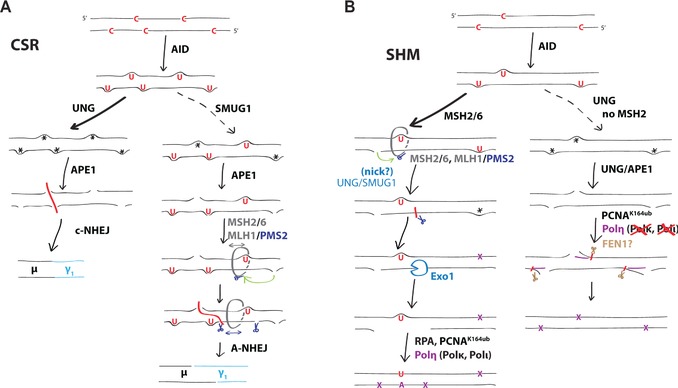
Excision of uracil introduced into the immunoglobulin loci by AID is central to antibody diversification. While predominantly carried out by the UNG uracil-DNA glycosylase as reflected by deficiency in immunoglobulin class switching in Ung(-/-) mice, the deficiency is incomplete, as evidenced by the emergence of switched IgG in the serum of Ung(-/-) mice. Lack of switching in mice deficient in both UNG and MSH2 suggested that mismatch repair initiated a backup pathway. We now show that most of the residual class switching in Ung(-/-) mice depends upon the endogenous SMUG1 uracil-DNA glycosylase, with in vitro switching to IgG1 as well as serum IgG3, IgG2b, and IgA greatly diminished in Ung(-/-) Smug1(-/-) mice, and that Smug1 partially compensates for Ung deficiency over time. Nonetheless, using a highly MSH2-dependent mechanism, Ung(-/-) Smug1(-/-) mice can still produce detectable levels of switched isotypes, especially IgG1. While not affecting the pattern of base substitutions, SMUG1 deficiency in an Ung(-/-) background further reduces somatic hypermutation at A:T base pairs. Our data reveal an essential requirement for uracil excision in class switching and in facilitating noncanonical mismatch repair for the A:T phase of hypermutation presumably by creating nicks near the U:G lesion recognized by MSH2.
Keywords: Class switching; DNA deamination; Somatic hypermutation; Uracil.
© 2014 The Authors. European Journal of Immunology published by WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures
Comment in
-
Refining the Neuberger model: Uracil processing by activated B cells.Eur J Immunol. 2014 Jul;44(7):1913-6. doi: 10.1002/eji.201444813. Epub 2014 Jun 30. Eur J Immunol. 2014. PMID: 24920531 Free PMC article.
References
-
- Honjo, T., Kinoshita, K. and Muramatsu, M., Molecular mechanism of class switch recombination: linkage with somatic hypermutation. Annu. Rev. Immunol. 2002. 20:165–196 - PubMed
-
- Di Noia, J. M. and Neuberger, M. S., Molecular mechanisms of antibody somatic hypermutation. Annu. Rev. Biochem. 2007. 76:1–22 - PubMed
-
- Rada, C., Ehrenstein, M. R., Neuberger, M. S. and Milstein, C., Hot spot focusing of somatic hypermutation in MSH2‐deficient mice suggests two stages of mutational targeting. Immunity 1998. 9:135–141 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
