

The genetics of microdeletion and microduplication syndromes: an update
- PMID: 24773319
- PMCID: PMC4476258
- DOI: 10.1146/annurev-genom-091212-153408
The genetics of microdeletion and microduplication syndromes: an update
Abstract
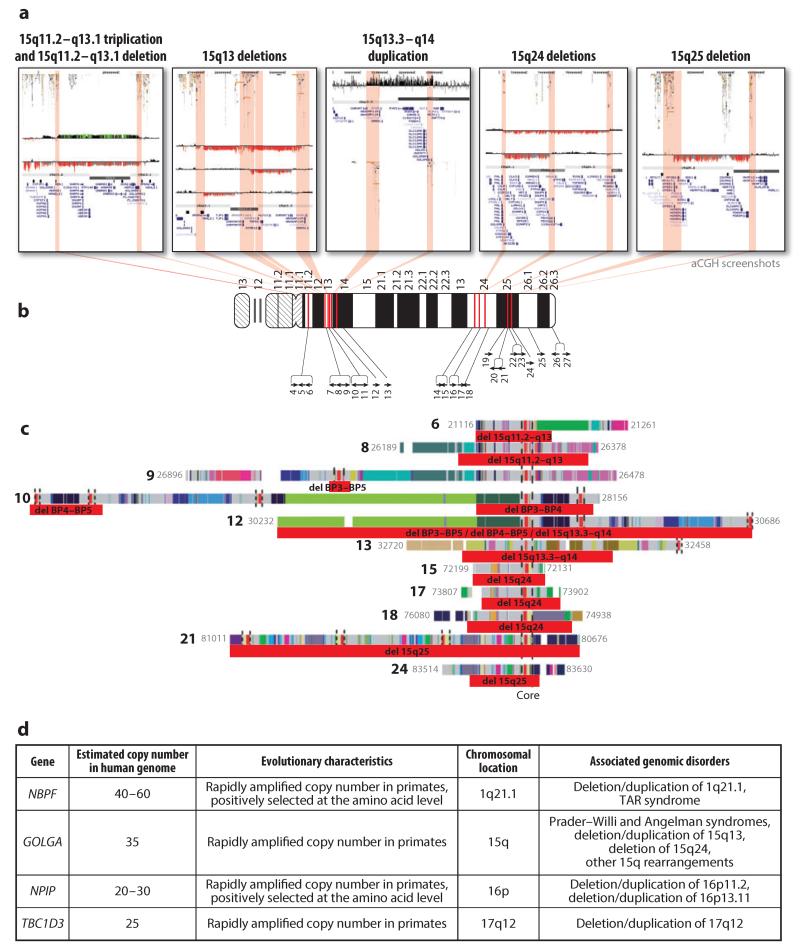
Chromosomal abnormalities, including microdeletions and microduplications, have long been associated with abnormal developmental outcomes. Early discoveries relied on a common clinical presentation and the ability to detect chromosomal abnormalities by standard karyotype analysis or specific assays such as fluorescence in situ hybridization. Over the past decade, the development of novel genomic technologies has allowed more comprehensive, unbiased discovery of microdeletions and microduplications throughout the human genome. The ability to quickly interrogate large cohorts using chromosome microarrays and, more recently, next-generation sequencing has led to the rapid discovery of novel microdeletions and microduplications associated with disease, including very rare but clinically significant rearrangements. In addition, the observation that some microdeletions are associated with risk for several neurodevelopmental disorders contributes to our understanding of shared genetic susceptibility for such disorders. Here, we review current knowledge of microdeletion/duplication syndromes, with a particular focus on recurrent rearrangement syndromes.
Keywords: copy-number variation; developmental delay; intellectual disability; microarray; nonallelic homologous recombination; recurrent rearrangement.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
