

Partitioning neuronal variability
- PMID: 24777419
- PMCID: PMC4135707
- DOI: 10.1038/nn.3711
Partitioning neuronal variability
Abstract
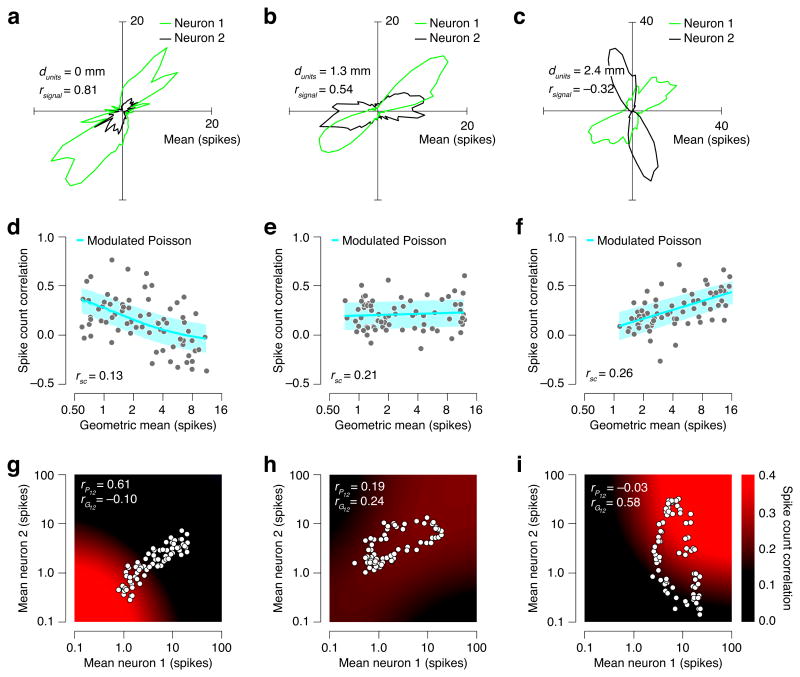
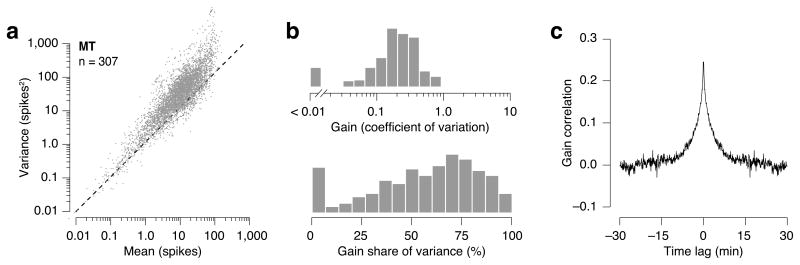
Responses of sensory neurons differ across repeated measurements. This variability is usually treated as stochasticity arising within neurons or neural circuits. However, some portion of the variability arises from fluctuations in excitability due to factors that are not purely sensory, such as arousal, attention and adaptation. To isolate these fluctuations, we developed a model in which spikes are generated by a Poisson process whose rate is the product of a drive that is sensory in origin and a gain summarizing stimulus-independent modulatory influences on excitability. This model provides an accurate account of response distributions of visual neurons in macaque lateral geniculate nucleus and cortical areas V1, V2 and MT, revealing that variability originates in large part from excitability fluctuations that are correlated over time and between neurons, and that increase in strength along the visual pathway. The model provides a parsimonious explanation for observed systematic dependencies of response variability and covariability on firing rate.
Figures





Comment in
-
Is there signal in the noise?Nat Neurosci. 2014 Jun;17(6):750-1. doi: 10.1038/nn.3722. Nat Neurosci. 2014. PMID: 24866037 No abstract available.
References
-
- Mainen ZF, Sejnowski TJ. Reliability of spike timing in neocortical neurons. Science. 1995;268:1503–1506. - PubMed
-
- Stevens CF, Zador A. When is an integrate-and-fire neuron like a Poisson neuron? In: Mozer M, Touretzky DS, Hasselmo M, editors. Advances in Neural Information Processing Systems. Vol. 8. 1996. pp. 103–109.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
