

Caveolin-1 regulates genomic action of the glucocorticoid receptor in neural stem cells
- PMID: 24777604
- PMCID: PMC4097667
- DOI: 10.1128/MCB.01121-13
Caveolin-1 regulates genomic action of the glucocorticoid receptor in neural stem cells
Abstract
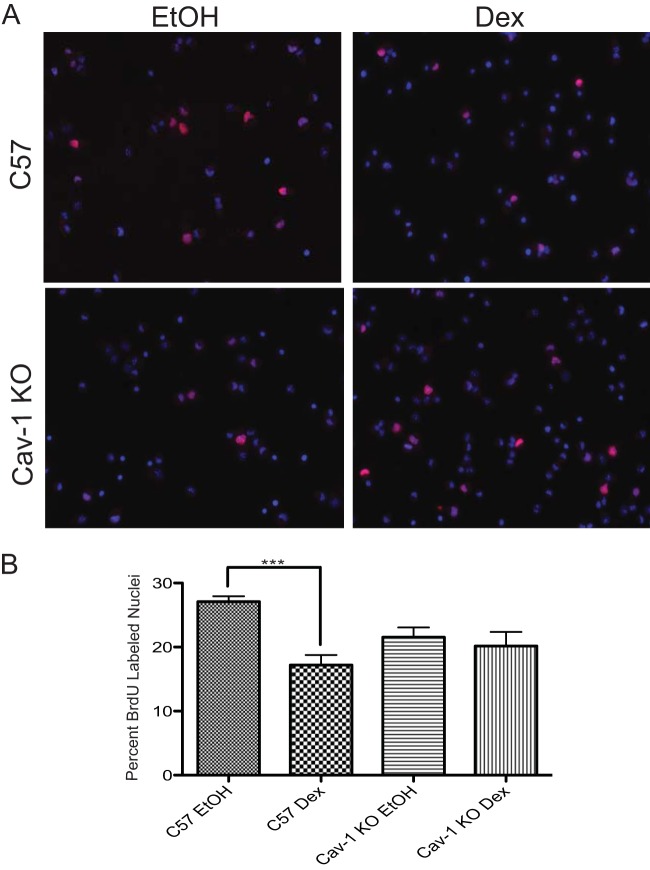
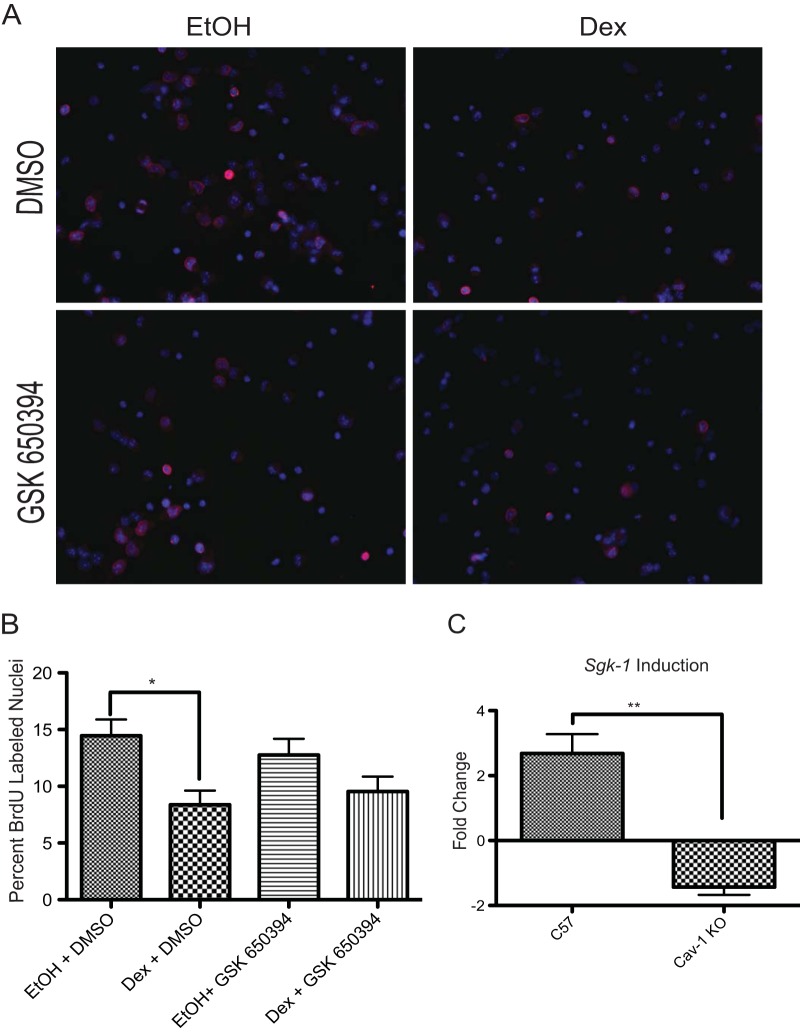
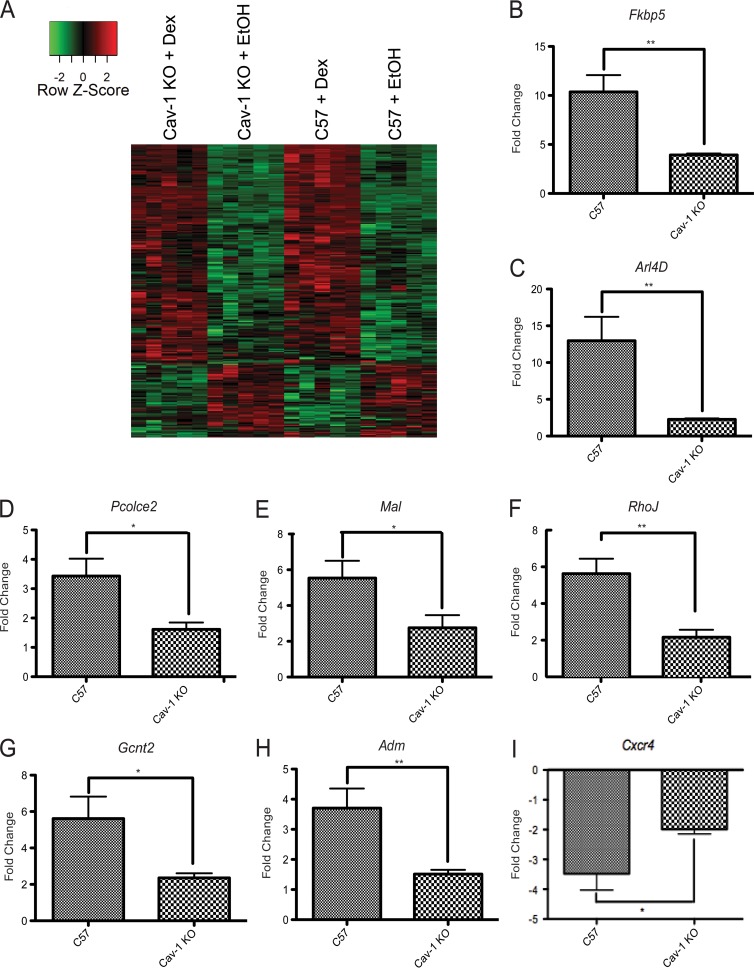
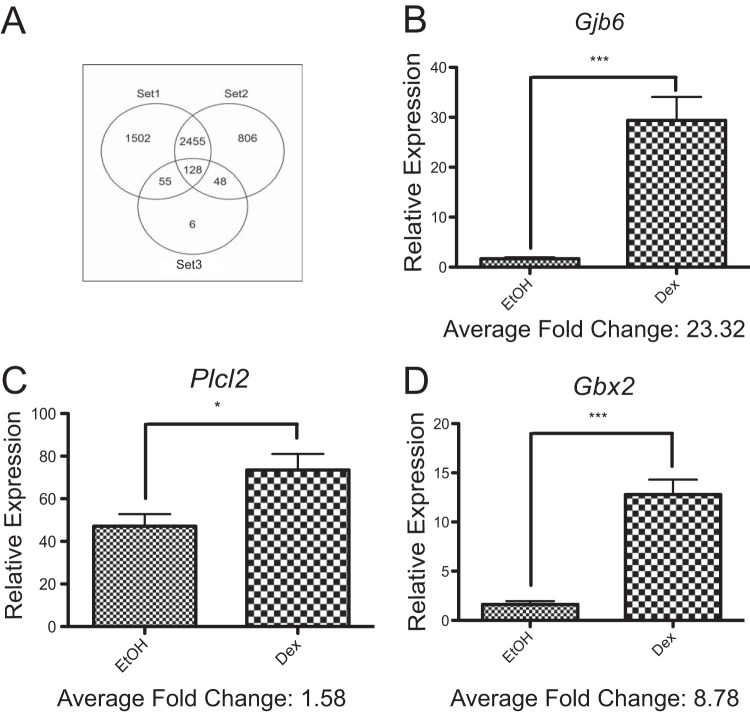
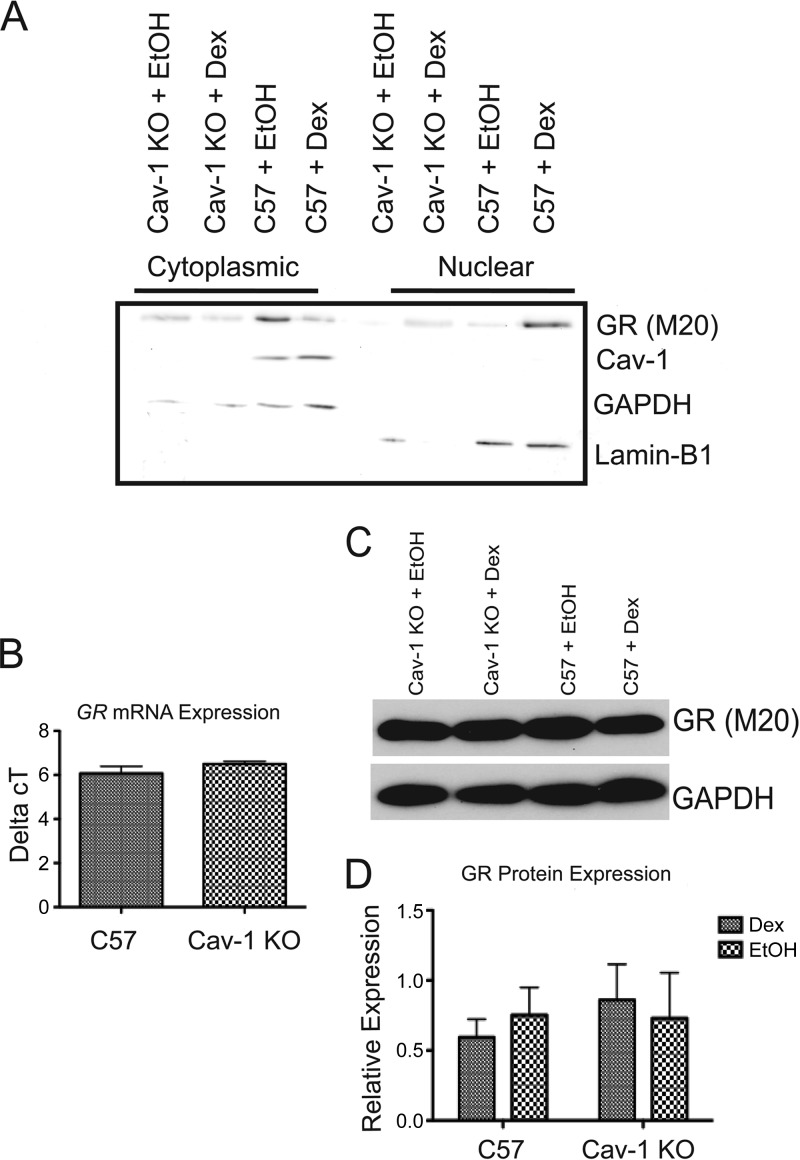
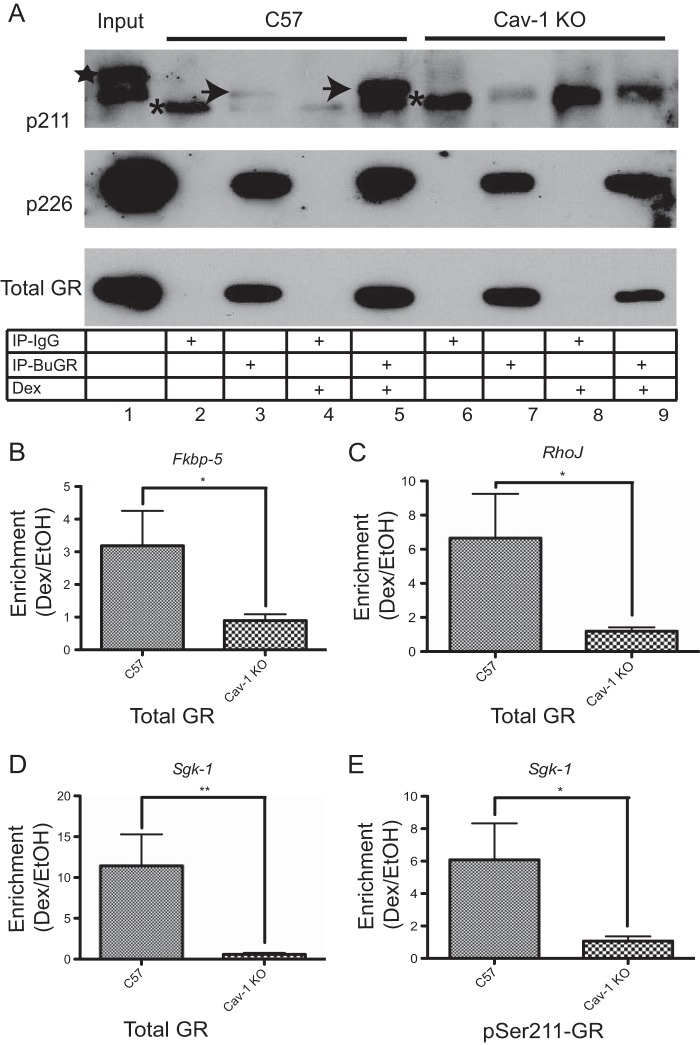
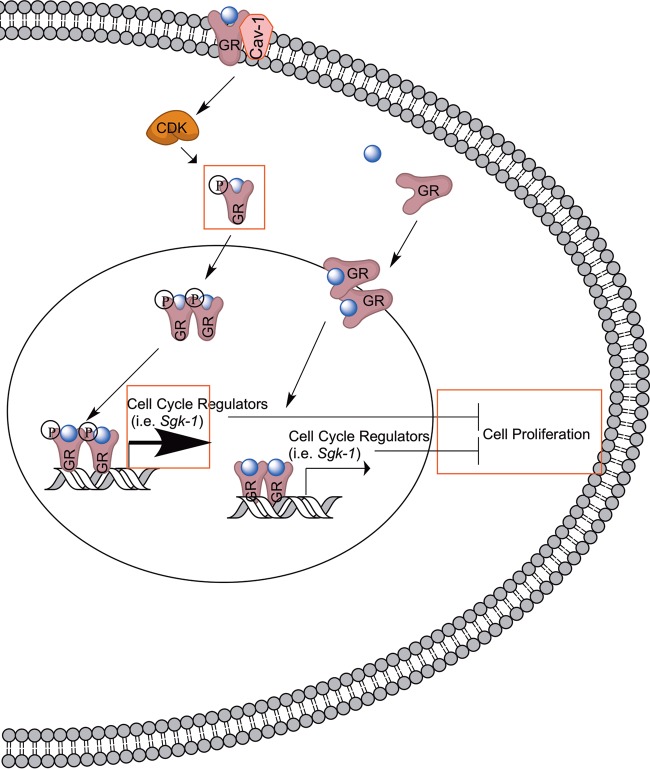
While glucocorticoids (GCs) are used clinically to treat many conditions, their neonatal and prenatal usage is increasingly controversial due to reports of delayed adverse outcomes, especially their effects on brain development. Such alterations may reflect the impact of GCs on neural progenitor/stem cell (NPSC) function. We previously demonstrated that the lipid raft protein caveolin-1 (Cav-1) was required for rapid GC signaling in embryonic mouse NPSCs operating through plasma membrane-bound glucocorticoid receptors (GRs). We show here that genomic GR signaling in NPSCs requires Cav-1. Loss of Cav-1 impacts the transcriptional response of many GR target genes (e.g., the serum- and glucocorticoid-regulated kinase 1 gene) that are likely to mediate the antiproliferative effects of GCs. Microarray analysis of wild-type C57 or Cav-1-deficient NPSCs identified approximately 100 genes that are differentially regulated by GC treatment. These changes in hormone responsiveness in Cav-1 knockout NPSCs are associated with the loss of GC-regulated phosphorylation of GR at serine 211 but not at serine 226. Chromatin recruitment of total GR to regulatory regions of target genes such as Fkbp-5, RhoJ, and Sgk-1, as well as p211-GR recruitment to Sgk-1, are compromised in Cav-1 knockout NPSCs. Cav-1 is therefore a multifunctional regulator of GR in NPSCs influencing both rapid and genomic action of the receptor to impact cell proliferation.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous