

The mirror mechanism: recent findings and perspectives
- PMID: 24778385
- PMCID: PMC4006191
- DOI: 10.1098/rstb.2013.0420
The mirror mechanism: recent findings and perspectives
Abstract
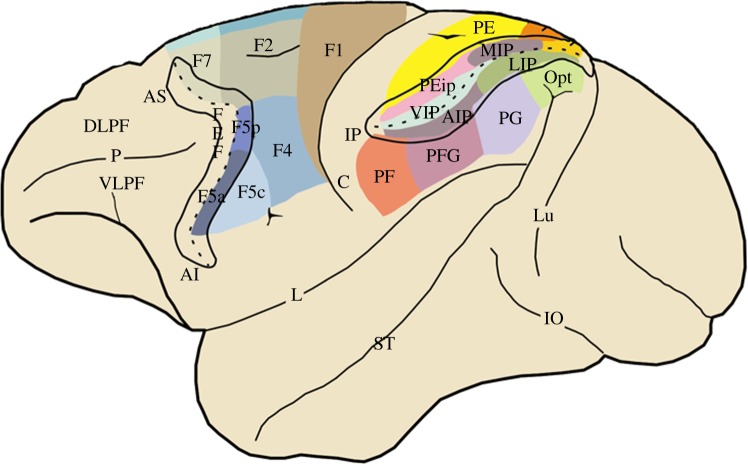
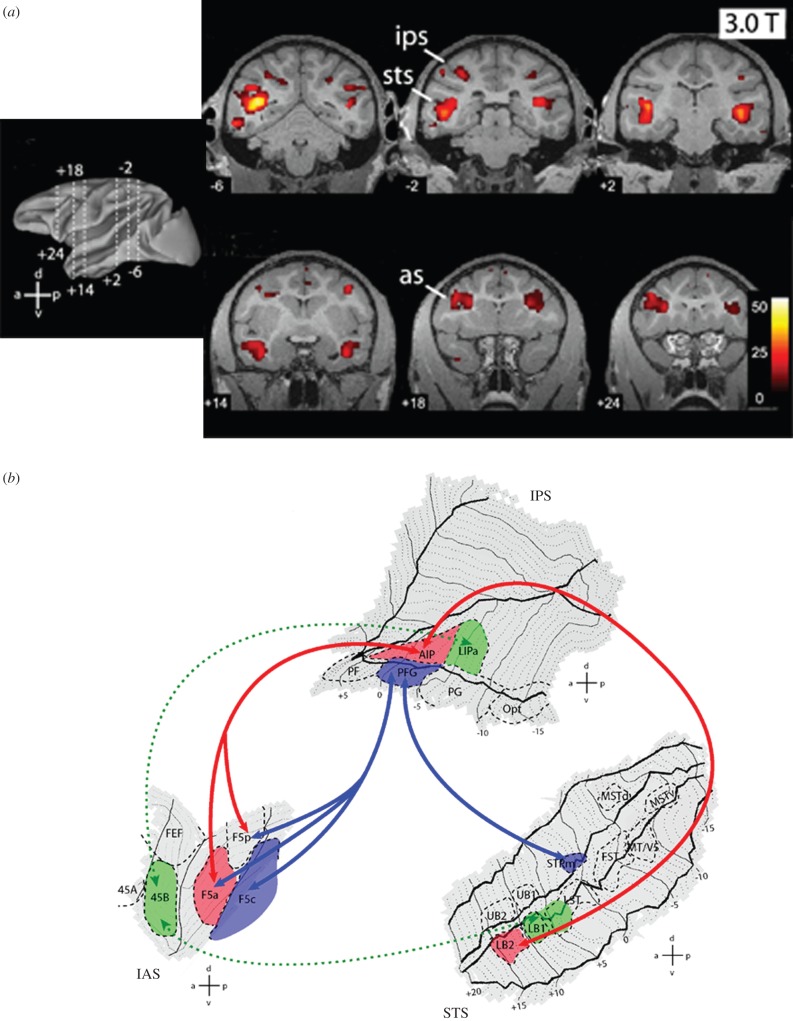
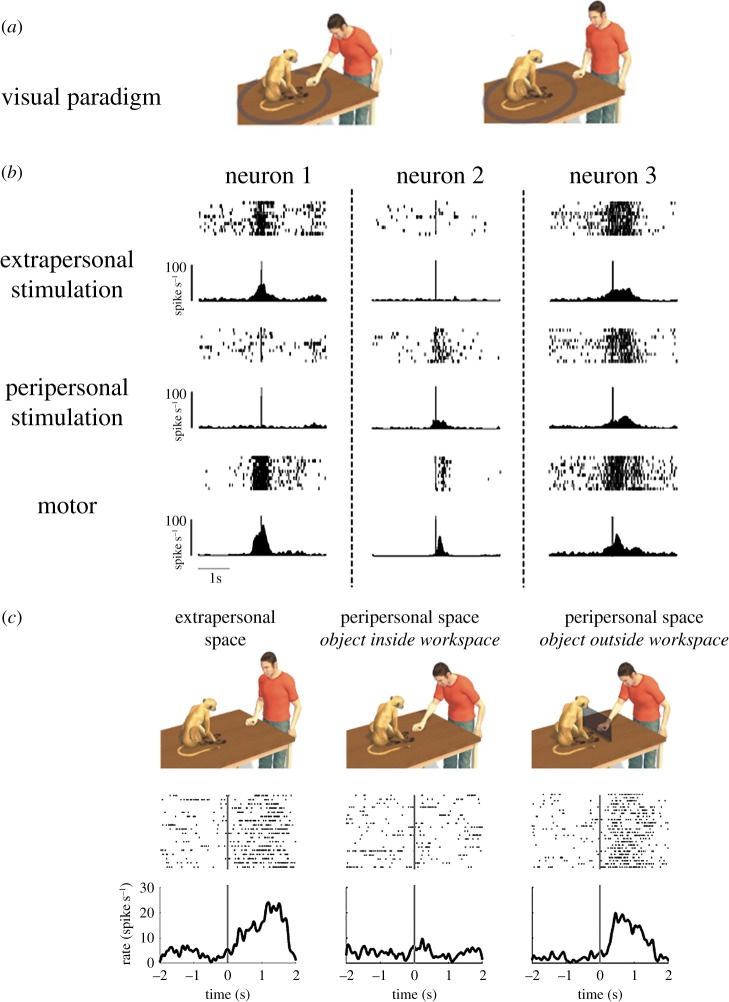
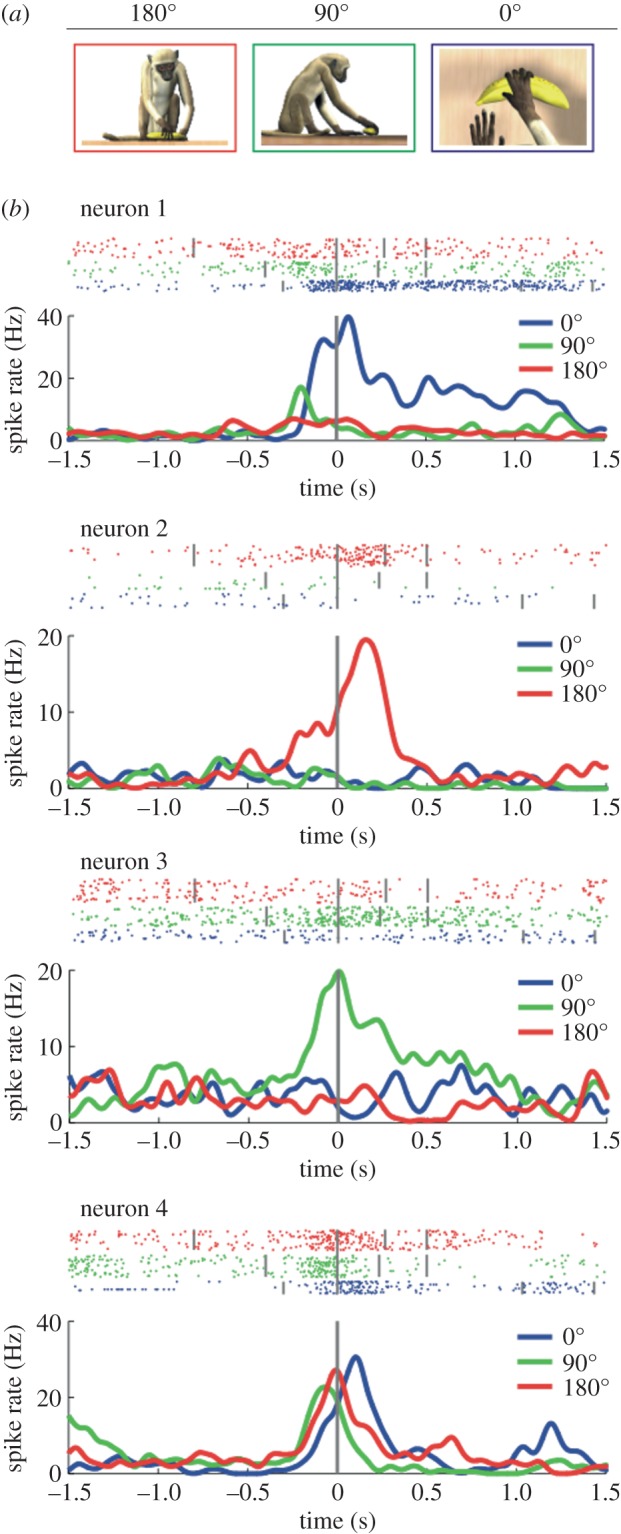
Mirror neurons are a specific type of visuomotor neuron that discharge both when a monkey executes a motor act and when it observes a similar motor act performed by another individual. In this article, we review first the basic properties of these neurons. We then describe visual features recently investigated which indicate that, besides encoding the goal of motor acts, mirror neurons are modulated by location in space of the observed motor acts, by the perspective from which the others' motor acts are seen, and by the value associated with the object on which others' motor acts are performed. In the last part of this article, we discuss the role of the mirror mechanism in planning actions and in understanding the intention underlying the others' motor acts. We also review some human studies suggesting that motor intention in humans may rely, as in the monkey, on the mirror mechanism.
Keywords: motor act; motor intention understanding; space-selective mirror neurons; subjective value; view-selective mirror neurons.
Figures
Similar articles
-
Cortical and subcortical connections of parietal and premotor nodes of the monkey hand mirror neuron network.Brain Struct Funct. 2018 May;223(4):1713-1729. doi: 10.1007/s00429-017-1582-0. Epub 2017 Dec 1. Brain Struct Funct. 2018. PMID: 29196811
-
Neurophysiological bases underlying the organization of intentional actions and the understanding of others' intention.Conscious Cogn. 2013 Sep;22(3):1095-104. doi: 10.1016/j.concog.2013.03.001. Epub 2013 Mar 30. Conscious Cogn. 2013. PMID: 23545395 Review.
-
Parietal lobe: from action organization to intention understanding.Science. 2005 Apr 29;308(5722):662-7. doi: 10.1126/science.1106138. Science. 2005. PMID: 15860620
-
Space-dependent representation of objects and other's action in monkey ventral premotor grasping neurons.J Neurosci. 2014 Mar 12;34(11):4108-19. doi: 10.1523/JNEUROSCI.4187-13.2014. J Neurosci. 2014. PMID: 24623789 Free PMC article.
-
Mirror neurons (and beyond) in the macaque brain: an overview of 20 years of research.Neurosci Lett. 2013 Apr 12;540:3-14. doi: 10.1016/j.neulet.2012.11.003. Epub 2012 Nov 13. Neurosci Lett. 2013. PMID: 23153827 Review.
Cited by
-
The Enactive Approach to Architectural Experience: A Neurophysiological Perspective on Embodiment, Motivation, and Affordances.Front Psychol. 2016 Mar 31;7:481. doi: 10.3389/fpsyg.2016.00481. eCollection 2016. Front Psychol. 2016. PMID: 27065937 Free PMC article.
-
Investigation of the Sense of Agency in Social Cognition, Based on Frameworks of Predictive Coding and Active Inference: A Simulation Study on Multimodal Imitative Interaction.Front Neurorobot. 2020 Sep 7;14:61. doi: 10.3389/fnbot.2020.00061. eCollection 2020. Front Neurorobot. 2020. PMID: 33013346 Free PMC article.
-
Cross-Modal Coordination of Face-Directed Gaze and Emotional Speech Production in School-Aged Children and Adolescents with ASD.Sci Rep. 2019 Dec 4;9(1):18301. doi: 10.1038/s41598-019-54587-z. Sci Rep. 2019. PMID: 31797950 Free PMC article.
-
The effect of interbrain synchronization in gesture observation: A fNIRS study.Brain Behav. 2020 Jul;10(7):e01663. doi: 10.1002/brb3.1663. Epub 2020 May 29. Brain Behav. 2020. PMID: 32469153 Free PMC article.
-
Mindreading in the balance: adults' mediolateral leaning and anticipatory looking foretell others' action preparation in a false-belief interactive task.R Soc Open Sci. 2020 Jan 29;7(1):191167. doi: 10.1098/rsos.191167. eCollection 2020 Jan. R Soc Open Sci. 2020. PMID: 32218946 Free PMC article.
References
-
- Rozzi S, Ferrari PF, Bonini L, Rizzolatti G, Fogassi L. 2008. Functional organization of inferior parietal lobule convexity in the macaque monkey: electrophysiological characterization of motor, sensory and mirror responses and their correlation with cytoarchitectonic areas. Eur. J. Neurosci. 28, 1569–1588. (10.1111/j.1460-9568.2008.06395.x) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
