

Longitudinal requirement for CD4+ T cell help for adenovirus vector-elicited CD8+ T cell responses
- PMID: 24778441
- PMCID: PMC4025612
- DOI: 10.4049/jimmunol.1302806
Longitudinal requirement for CD4+ T cell help for adenovirus vector-elicited CD8+ T cell responses
Abstract
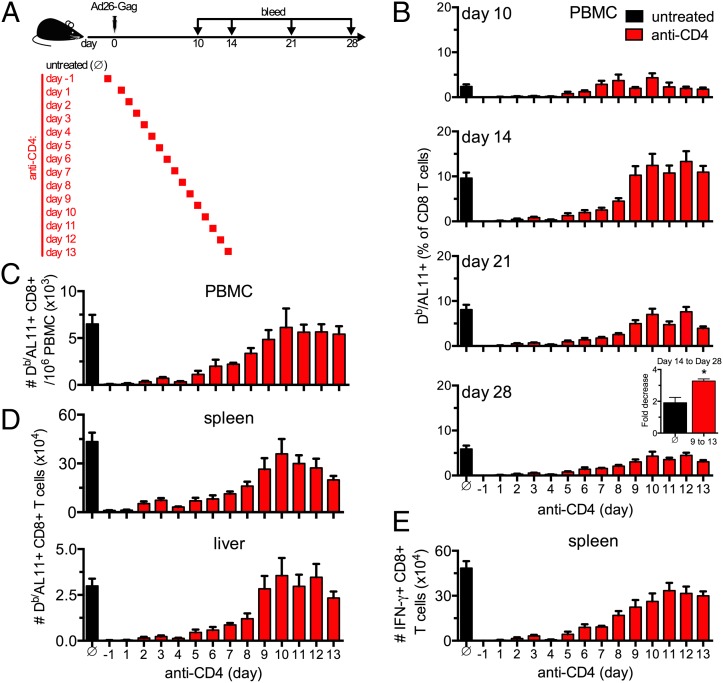
Despite the widespread use of replication-incompetent recombinant adenovirus (Ad) vectors as candidate vaccine platforms, the mechanism by which these vectors elicit CD8(+) T cell responses remains poorly understood. Our data demonstrate that induction and maintenance of CD8(+) T cell responses by Ad vector immunization is longitudinally dependent on CD4(+) T cell help for a prolonged period. Depletion of CD4(+) T cells in wild type mice within the first 8 d following Ad immunization resulted in dramatically reduced induction of Ag-specific CD8(+) T cells, decreased T-bet and eomesodermin expression, impaired KLRG1(+) effector differentiation, and atypical expression of the memory markers CD127, CD27, and CD62L. Moreover, these CD8(+) T cells failed to protect against a lethal recombinant Listeria monocytogenes challenge. Depletion of CD4(+) T cells between weeks 1 and 4 following immunization resulted in increased contraction of memory CD8(+) T cells. These data demonstrate a prolonged temporal requirement for CD4(+) T cell help for vaccine-elicited CD8(+) T cell responses in mice. These findings have important implications in the design of vaccines aimed at eliciting CD8(+) T cell responses and may provide insight into the impaired immunogenicity of vaccines in the context of AIDS and other CD4(+) T cell immune deficiencies.
Copyright © 2014 by The American Association of Immunologists, Inc.
Figures








Similar articles
-
Depletion of CD4+ T cells during immunization with nonviable Listeria monocytogenes causes enhanced CD8+ T cell-mediated protection against listeriosis.J Immunol. 2004 Mar 1;172(5):3167-72. doi: 10.4049/jimmunol.172.5.3167. J Immunol. 2004. PMID: 14978123
-
A specific role for B cells in the generation of CD8 T cell memory by recombinant Listeria monocytogenes.J Immunol. 2003 Feb 1;170(3):1443-51. doi: 10.4049/jimmunol.170.3.1443. J Immunol. 2003. PMID: 12538706
-
Adenovirus-based vaccine against Listeria monocytogenes: extending the concept of invariant chain linkage.J Immunol. 2013 Oct 15;191(8):4152-64. doi: 10.4049/jimmunol.1301290. Epub 2013 Sep 16. J Immunol. 2013. PMID: 24043891
-
Engineered Listeria monocytogenes as an AIDS vaccine.Vaccine. 2002 May 6;20(15):2007-10. doi: 10.1016/s0264-410x(02)00088-9. Vaccine. 2002. PMID: 11983264 Review.
-
Listeria monocytogenes: a live vector able to deliver heterologous protein within the cytosol and to drive a CD8 dependent T cell response.Biologicals. 1995 Jun;23(2):135-43. doi: 10.1006/biol.1995.0024. Biologicals. 1995. PMID: 7546656 Review.
Cited by
-
The differences in immunoadjuvant mechanisms of TLR3 and TLR4 agonists on the level of antigen-presenting cells during immunization with recombinant adenovirus vector.BMC Immunol. 2018 Jul 28;19(1):26. doi: 10.1186/s12865-018-0264-x. BMC Immunol. 2018. PMID: 30055563 Free PMC article.
-
Immunogenicity and Cross-Reactivity of Rhesus Adenoviral Vectors.J Virol. 2018 May 14;92(11):e00159-18. doi: 10.1128/JVI.00159-18. Print 2018 Jun 1. J Virol. 2018. PMID: 29563285 Free PMC article.
-
Immediate Dysfunction of Vaccine-Elicited CD8+ T Cells Primed in the Absence of CD4+ T Cells.J Immunol. 2016 Sep 1;197(5):1809-22. doi: 10.4049/jimmunol.1600591. Epub 2016 Jul 22. J Immunol. 2016. PMID: 27448585 Free PMC article.
-
Novel Concepts for HIV Vaccine Vector Design.mSphere. 2017 Dec 6;2(6):e00415-17. doi: 10.1128/mSphere.00415-17. eCollection 2017 Nov-Dec. mSphere. 2017. PMID: 29242831 Free PMC article. Review.
-
Distinct CD4 T-cell effects on primary versus recall CD8 T-cell responses during viral encephalomyelitis.Immunology. 2015 Mar;144(3):374-386. doi: 10.1111/imm.12378. Immunology. 2015. PMID: 25187405 Free PMC article.
References
-
- Sullivan N. J., Hensley L., Asiedu C., Geisbert T. W., Stanley D., Johnson J., Honko A., Olinger G., Bailey M., Geisbert J. B., et al. 2011. CD8+ cellular immunity mediates rAd5 vaccine protection against Ebola virus infection of nonhuman primates. Nat. Med. 17: 1128–1131 - PubMed
-
- Alexander J., Ward S., Mendy J., Manayani D. J., Farness P., Avanzini J. B., Guenther B., Garduno F., Jow L., Snarsky V., et al. 2012. Pre-clinical evaluation of a replication-competent recombinant adenovirus serotype 4 vaccine expressing influenza H5 hemagglutinin. PLoS ONE 7: e31177. - PMC - PubMed
-
- Colloca S., Barnes E., Folgori A., Ammendola V., Capone S., Cirillo A., Siani L., Naddeo M., Grazioli F., Esposito M. L., et al. 2012. Vaccine vectors derived from a large collection of simian adenoviruses induce potent cellular immunity across multiple species. Sci. Transl. Med. 4: 115ra112 - PMC - PubMed
-
- Xiang Z. Q., Yang Y., Wilson J. M., Ertl H. C. 1996. A replication-defective human adenovirus recombinant serves as a highly efficacious vaccine carrier. Virology 219: 220–227 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
