

Endothelial PINK1 mediates the protective effects of NLRP3 deficiency during lethal oxidant injury
- PMID: 24778451
- PMCID: PMC4047670
- DOI: 10.4049/jimmunol.1400653
Endothelial PINK1 mediates the protective effects of NLRP3 deficiency during lethal oxidant injury
Abstract
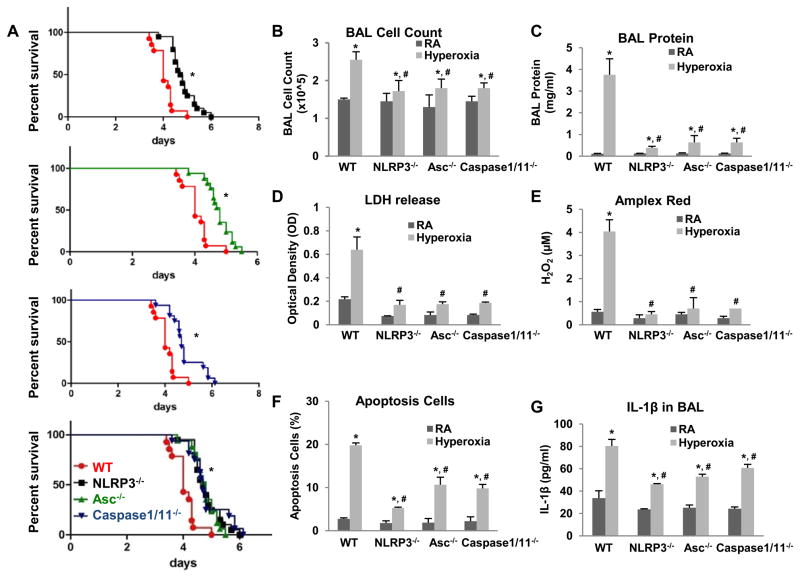
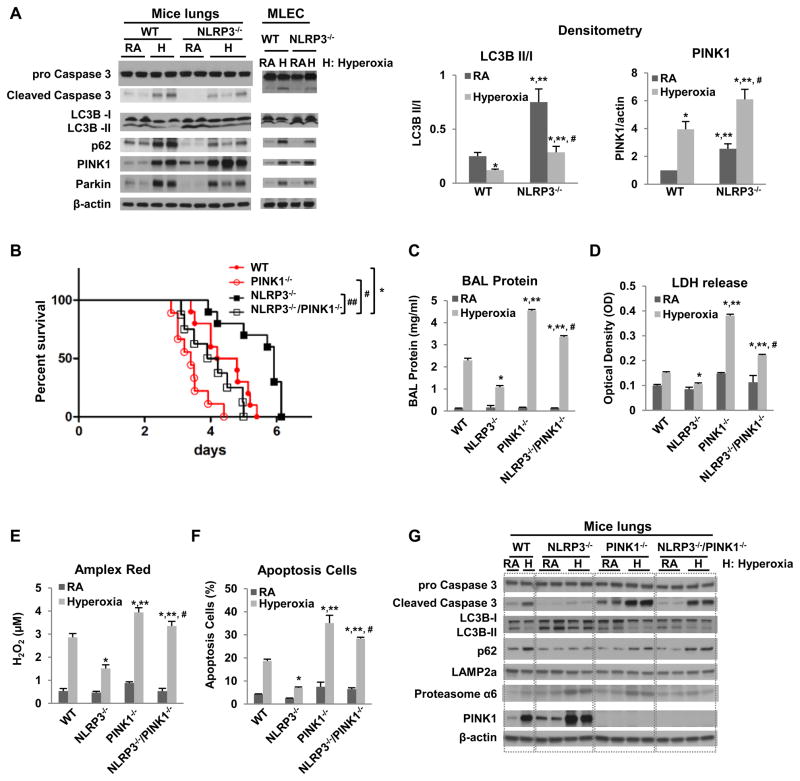
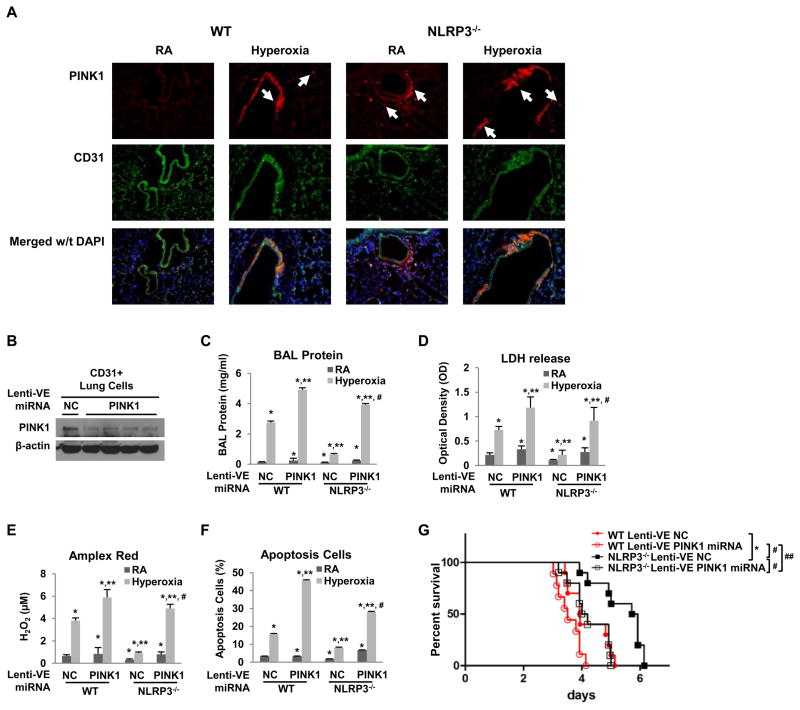
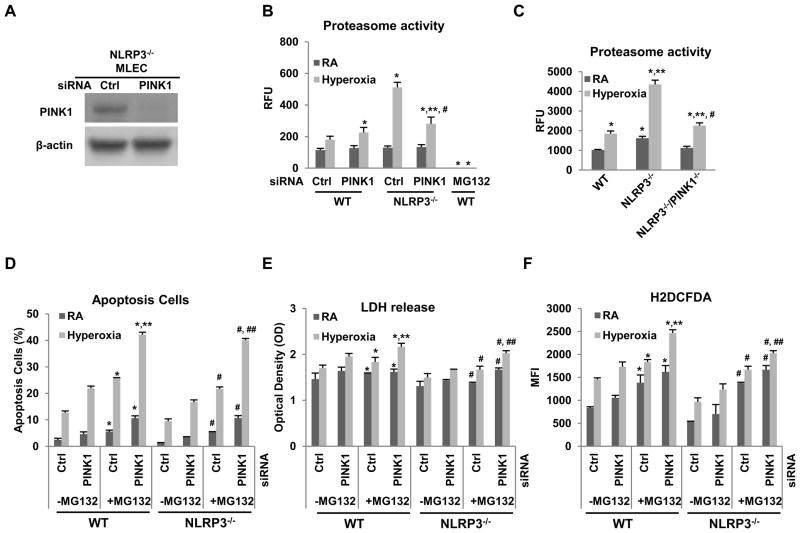
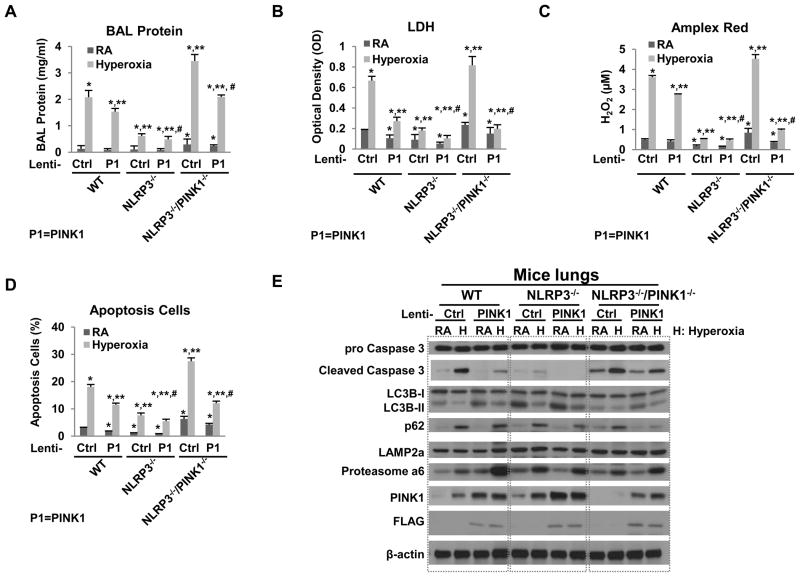
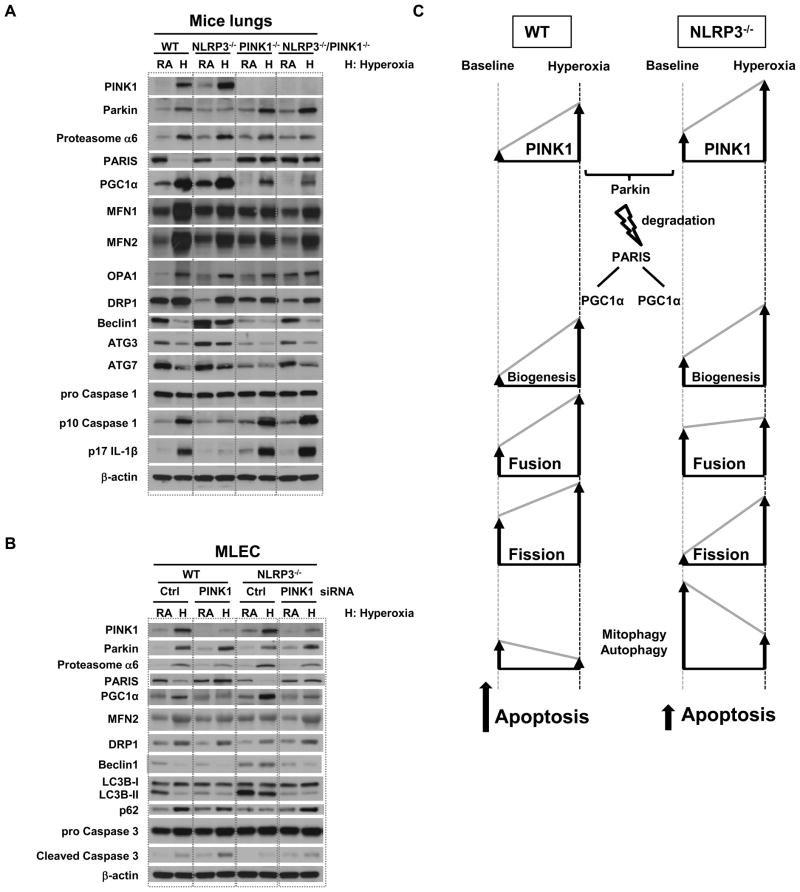
High levels of inspired oxygen, hyperoxia, are frequently used in patients with acute respiratory failure. Hyperoxia can exacerbate acute respiratory failure, which has high mortality and no specific therapies. We identified novel roles for PTEN-induced putative kinase 1 (PINK1), a mitochondrial protein, and the cytosolic innate immune protein NLRP3 in the lung and endothelium. We generated double knockouts (PINK1(-/-)/NLRP3(-/-)), as well as cell-targeted PINK1 silencing and lung-targeted overexpression constructs, to specifically show that PINK1 mediates cytoprotection in wild-type and NLRP3(-/-) mice. The ability to resist hyperoxia is proportional to PINK1 expression. PINK1(-/-) mice were the most susceptible; wild-type mice, which induced PINK1 after hyperoxia, had intermediate susceptibility; and NLRP3(-/-) mice, which had high basal and hyperoxia-induced PINK1, were the least susceptible. Genetic deletion of PINK1 or PINK1 silencing in the lung endothelium increased susceptibility to hyperoxia via alterations in autophagy/mitophagy, proteasome activation, apoptosis, and oxidant generation.
Copyright © 2014 by The American Association of Immunologists, Inc.
Figures
References
-
- Fisher AB. Oxygen therapy. Side effects and toxicity. Am Rev Respir Dis. 1980;122:61–69. - PubMed
-
- Sutterwala FS, Ogura Y, Szczepanik M, Lara-Tejero M, Lichtenberger GS, Grant EP, Bertin J, Coyle AJ, Galan JE, Askenase PW, Flavell RA. Critical role for NALP3/CIAS1/Cryopyrin in innate and adaptive immunity through its regulation of caspase-1. Immunity. 2006;24:317–327. - PubMed
-
- Kuida K, Lippke JA, Ku G, Harding MW, Livingston DJ, Su MS, Flavell RA. Altered cytokine export and apoptosis in mice deficient in interleukin-1 beta converting enzyme. Science. 1995;267:2000–2003. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
