

Silencing VDAC1 Expression by siRNA Inhibits Cancer Cell Proliferation and Tumor Growth In Vivo
- PMID: 24781191
- PMCID: PMC4011124
- DOI: 10.1038/mtna.2014.9
Silencing VDAC1 Expression by siRNA Inhibits Cancer Cell Proliferation and Tumor Growth In Vivo
Erratum in
-
Silencing VDAC1 Expression by siRNA Inhibits Cancer Cell Proliferation and Tumor Growth In Vivo.Mol Ther Nucleic Acids. 2017 Sep 15;8:493. doi: 10.1016/j.omtn.2017.08.008. Epub 2017 Aug 19. Mol Ther Nucleic Acids. 2017. PMID: 28918049 Free PMC article. No abstract available.
Abstract
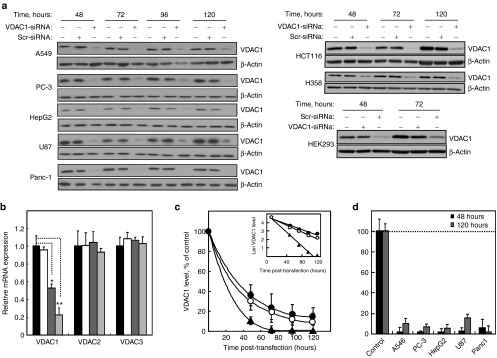
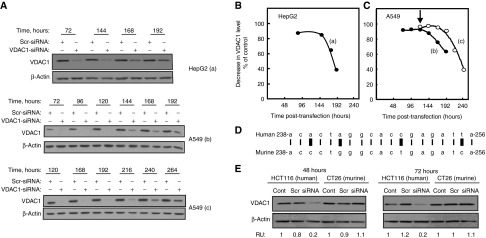
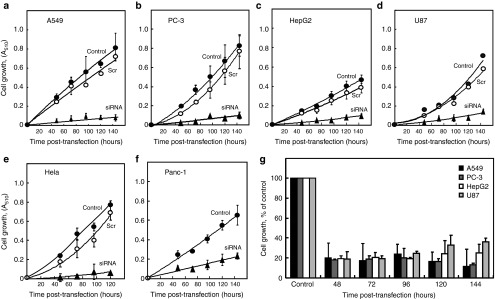
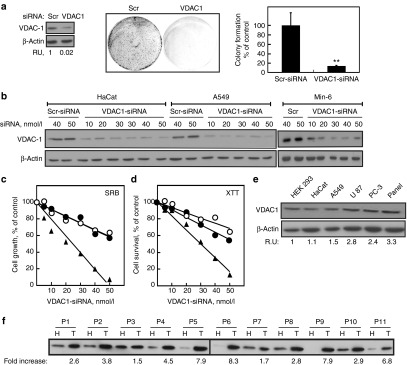
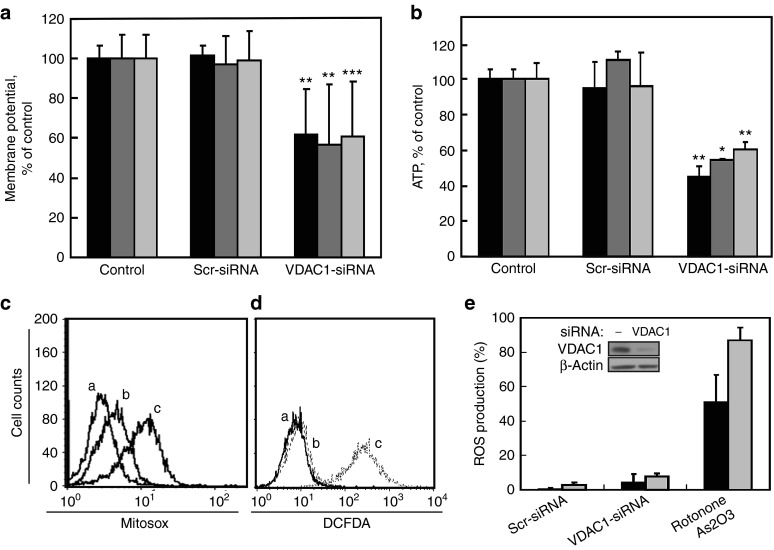
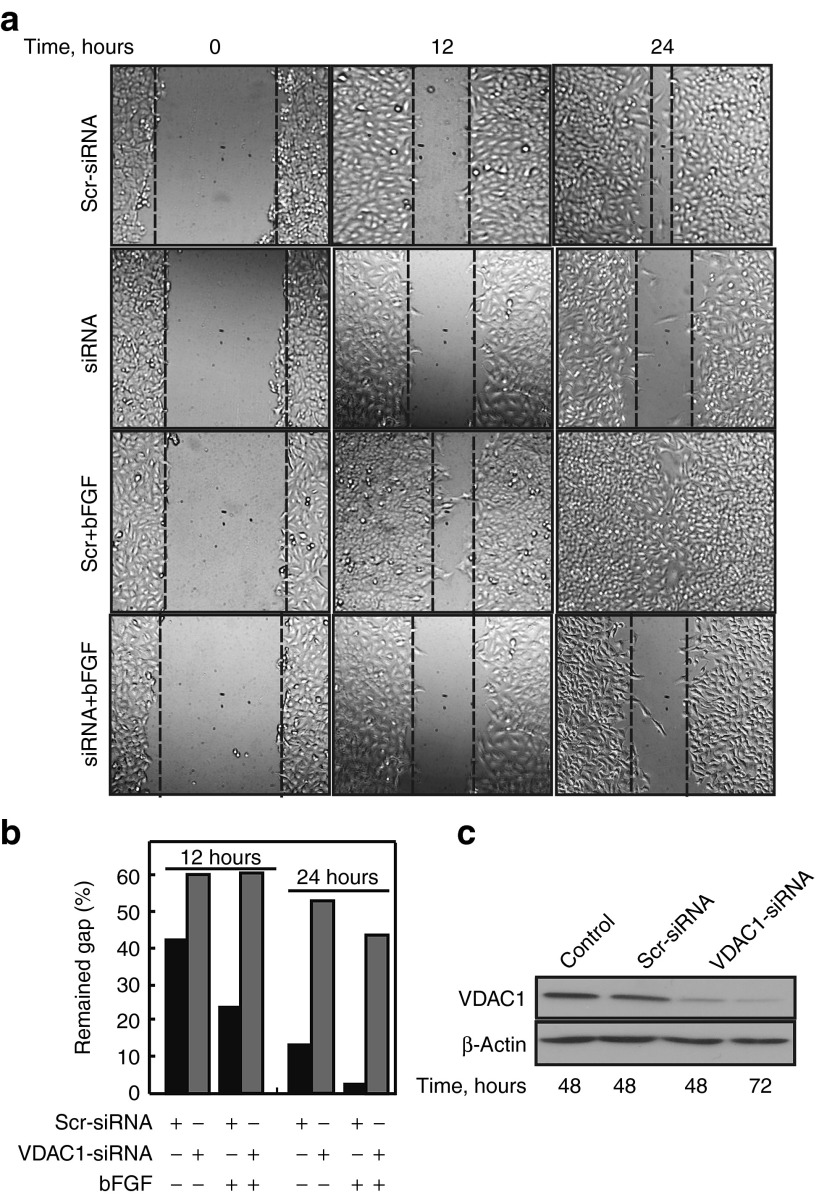
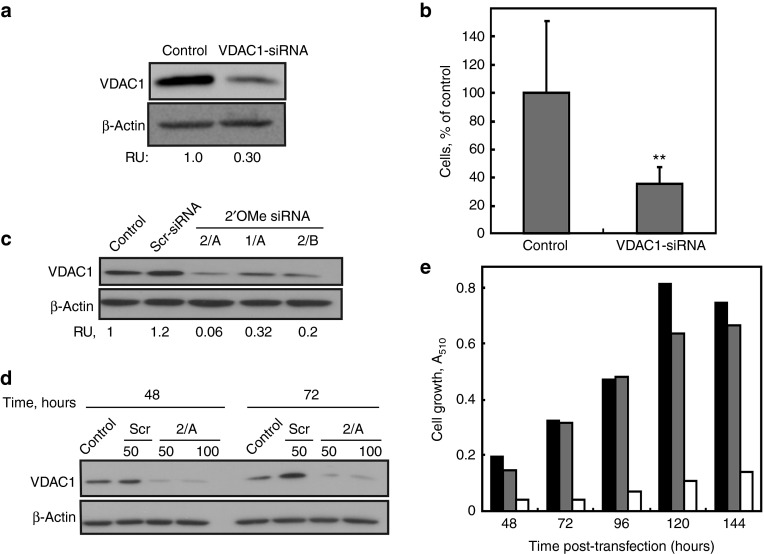
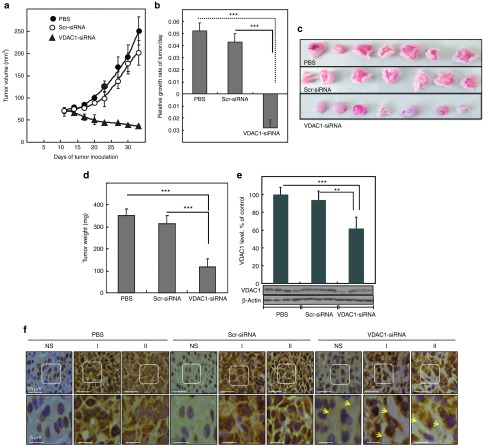
Alterations in cellular metabolism and bioenergetics are vital for cancer cell growth and motility. Here, the role of the mitochondrial protein voltage-dependent anion channel (VDAC1), a master gatekeeper regulating the flux of metabolites and ions between mitochondria and the cytoplasm, in regulating the growth of several cancer cell lines was investigated by silencing VDAC1 expression using small interfering RNA (siRNA). A single siRNA specific to the human VDAC1 sequence at nanomolar concentrations led to some 90% decrease in VDAC1 levels in the lung A549 and H358, prostate PC-3, colon HCT116, glioblastoma U87, liver HepG2, and pancreas Panc-1 cancer cell lines. VDAC1 silencing persisted 144 hours post-transfection and resulted in profound inhibition of cell growth in cancer but not in noncancerous cells, with up to 90% inhibition being observed over 5 days that was prolonged by a second transfection. Cells expressing low VDAC1 levels showed decreased mitochondrial membrane potential and adenoside triphosphate (ATP) levels, suggesting limited metabolite exchange between mitochondria and cytosol. Moreover, cells silenced for VDAC1 expression showed decreased migration, even in the presence of the wound healing accelerator basic fibroblast growth factor (bFGF). VDAC1-siRNA inhibited cancer cell growth in a Matrigel-based assay in host nude mice. Finally, in a xenograft lung cancer mouse model, chemically modified VDAC1-siRNA not only inhibited tumor growth but also resulted in tumor regression. This study thus shows that VDAC1 silencing by means of RNA interference (RNAi) dramatically inhibits cancer cell growth and tumor development by disabling the abnormal metabolic behavior of cancer cells, potentially paving the way for a more effective pipeline of anticancer drugs.
Figures
References
-
- Petrocca F, Lieberman J. Promise and challenge of RNA interference-based therapy for cancer. J Clin Oncol. 2011;29:747–754. - PubMed
-
- Soutschek J, Akinc A, Bramlage B, Charisse K, Constien R, Donoghue M, et al. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature. 2004;432:173–178. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
