

G-protein stimulatory subunit alpha and Gq/11α G-proteins are both required to maintain quiescent stem-like chondrocytes
- PMID: 24781502
- PMCID: PMC4154354
- DOI: 10.1038/ncomms4673
G-protein stimulatory subunit alpha and Gq/11α G-proteins are both required to maintain quiescent stem-like chondrocytes
Abstract
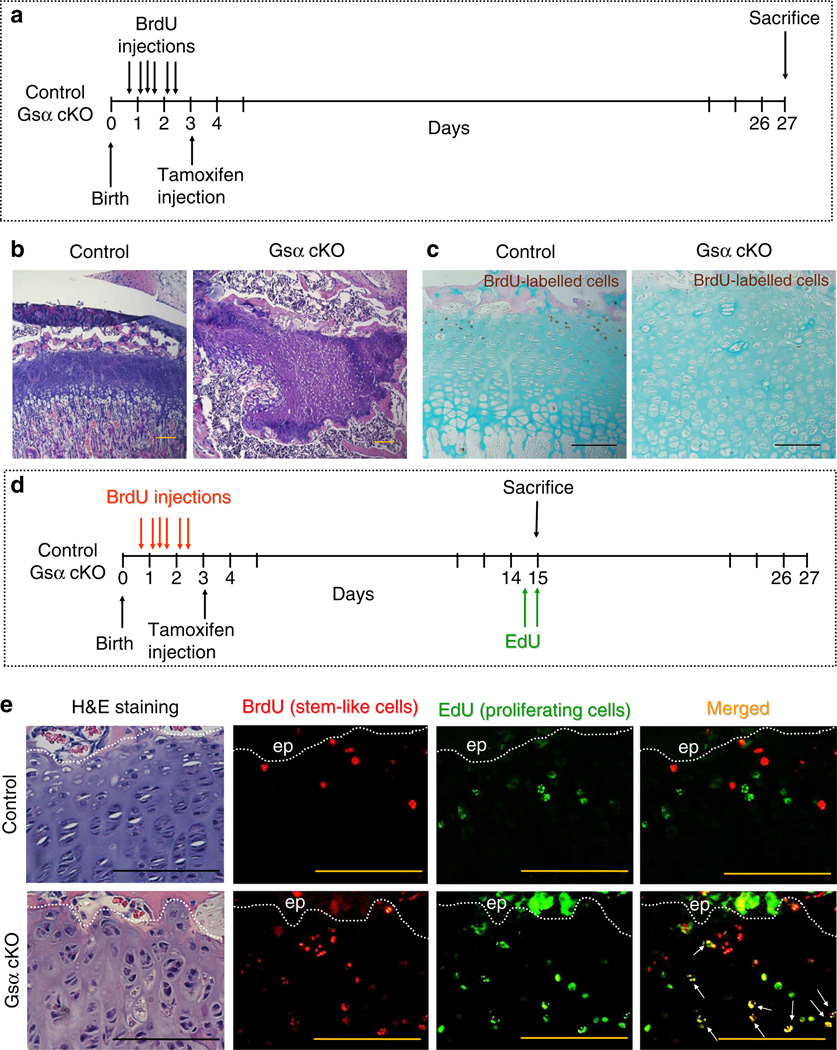
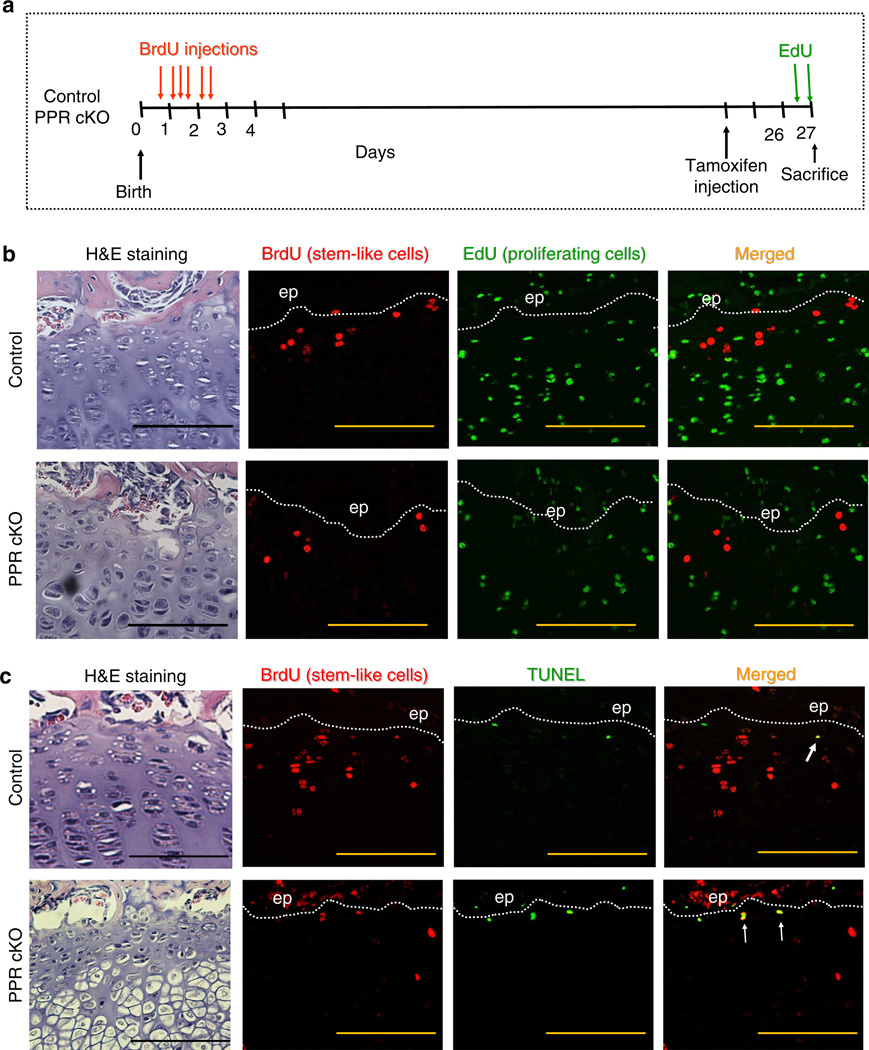
Round chondrocytes in the resting zone of the growth plate provide precursors for columnar chondrocytes and have stem-like properties. Here we demonstrate that these stem-like chondrocytes undergo apoptosis in the absence of the receptor (PPR) for parathyroid hormone-related protein. We examine the possible roles of heterotrimeric G-proteins activated by the PPR. Inactivation of the G-protein stimulatory α-subunit (G(s)α) leads to accelerated differentiation of columnar chondrocytes, as seen in the PPR knockout, but a remnant of growth cartilage remains, in contrast to disappearance of the growth cartilage in the PPR knockout. Stem-like chondrocytes lose their quiescence and proliferate upon G(s)α ablation. Inactivation of G(s)α in mice with a mutant PPR that cannot activate G proteins, Gq and G11, leads to a PPR knockout-like phenotype. Thus, G(s)α is the major mediator of the anti-differentiation action of the PPR, while activation of both G(s)α and Gq/11α is required for quiescence of stem-like chondrocytes.
Figures








References
-
- Kember NF, Lambert BE. Slowly cycling cells in growing bone. Cell Tissue Kinet. 1981;14:327–330. - PubMed
-
- Kember NF. Cell division in endochondral ossification. A study of cell proliferation in rat bones by the method of tritiated thymidine autoradiography. J. Bone Joint Surg. Br. 1960;42B:824–839. - PubMed
-
- Abad V, et al. The role of the resting zone in growth plate chondrogenesis. Endocrinology. 2002;143:1851–1857. - PubMed
-
- Mackie EJ, Tatarczuch L, Mirams M. The growth plate chondrocyte and endochondral ossification. J. Endocrinol. 2011;211:109–121. - PubMed
-
- Kronenberg HM. Developmental regulation of the growth plate. Nature. 2003;423:332–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
