

Competence and regulatory interactions during regeneration in plants
- PMID: 24782880
- PMCID: PMC3990048
- DOI: 10.3389/fpls.2014.00142
Competence and regulatory interactions during regeneration in plants
Abstract
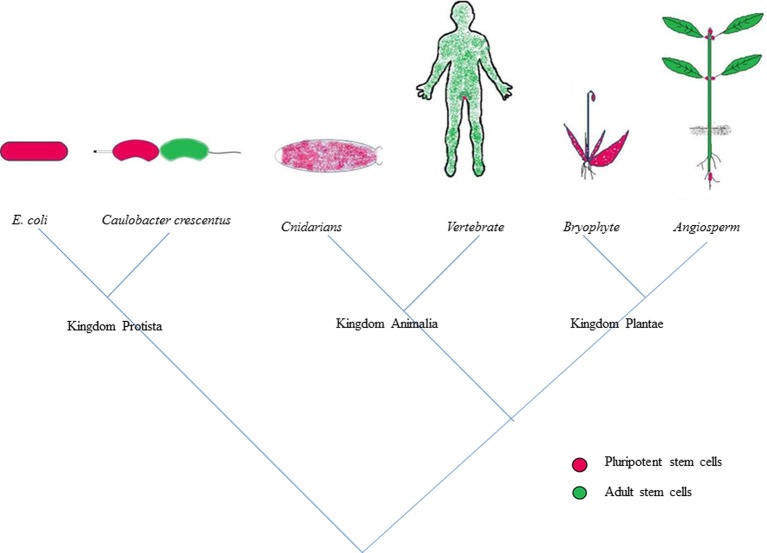
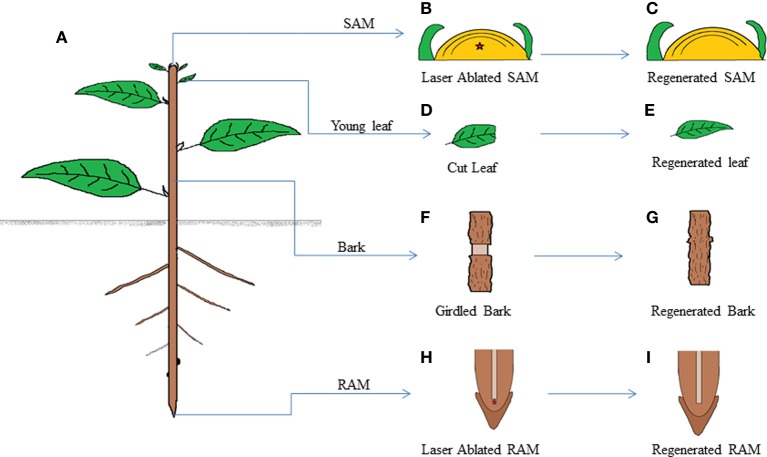
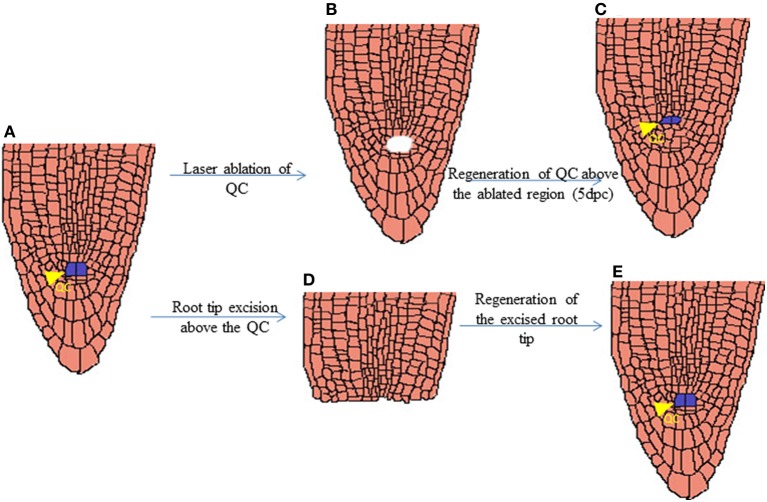
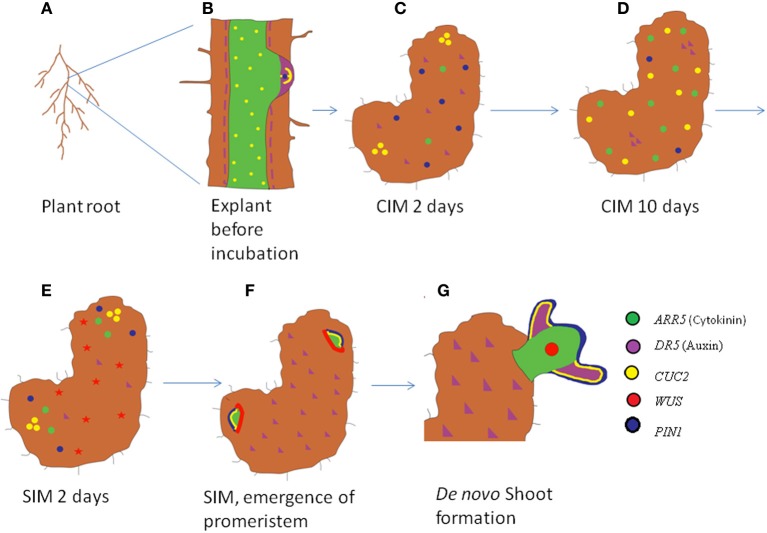
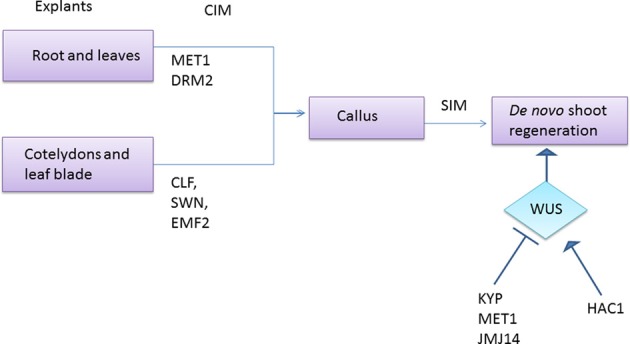
The ability to regenerate is widely exploited by multitudes of organisms ranging from unicellular bacteria to multicellular plants for their propagation and repair. But the levels of competence for regeneration vary from species to species. While variety of living cells of a plant display regeneration ability, only a few set of cells maintain their stemness in mammals. This highly pliable nature of plant cells in-terms of regeneration can be attributed to their high developmental plasticity. De novo organ initiation can be relatively easily achieved in plants by proper hormonal regulations. Elevated levels of plant hormone auxin induces the formation of proliferating mass of pluripotent cells called callus, which predominantly express lateral root meristem markers and hence is having an identity similar to lateral root primordia. Organ formation can be induced from the callus by modulating the ratio of hormones. An alternative for de novo organogenesis is by the forced expression of plant specific transcription factors. The mechanisms by which plant cells attain competence for regeneration on hormonal treatment or forced expression remain largely elusive. Recent studies have provided some insight into how the epigenetic modifications in plants affect this competence. In this review we discuss the present understanding of regenerative biology in plants and scrutinize the future prospectives of this topic. While discussing about the regeneration in the sporophyte of angiosperms which is well studied, here we outline the regenerative biology of the gametophytic phase and discuss about various strategies of regeneration that have evolved in the domain of life so that a common consensus on the entire process of regeneration can be made.
Keywords: De novo organ initiation; auxin; callus; competence for regeneration; cytokinin; epigenetic modifications.
Figures

References
-
- Abel S., Nguyen M. D., Chow W., Theologis A. (1995). ACS4, a primary indoleacetic acid-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis thaliana. Structural characterization, expression in Escherichia coli, and expression characteristics in response to auxin [corrected]. J. Biol. Chem. 270, 19093–19099 10.1074/jbc.270.32.19093 - DOI - PubMed
-
- Aguirre-Lipperheide M., Estrada-Rodríyuez F. J., Evans L. V. (1995). Facts, problems, and needs in seaweed tissue culture: an appraisal. J. Phycol. 31, 677–688 10.1111/j.0022-3646.1995.00677 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources