

Mössbauer, EPR, and modeling study of iron trafficking and regulation in Δccc1 and CCC1-up Saccharomyces cerevisiae
- PMID: 24785783
- PMCID: PMC4025571
- DOI: 10.1021/bi500002n
Mössbauer, EPR, and modeling study of iron trafficking and regulation in Δccc1 and CCC1-up Saccharomyces cerevisiae
Abstract
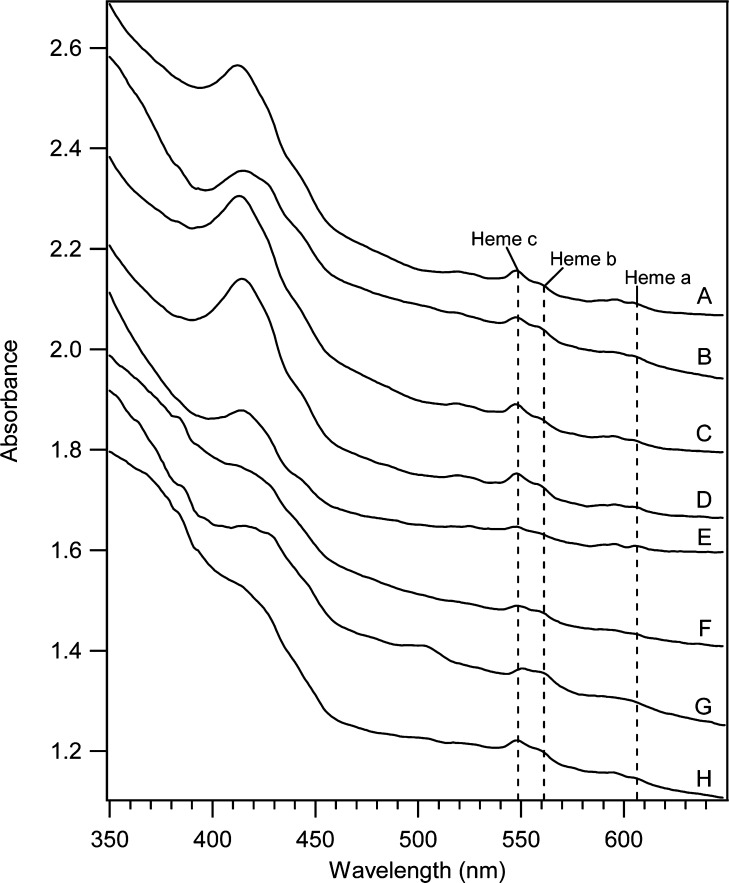
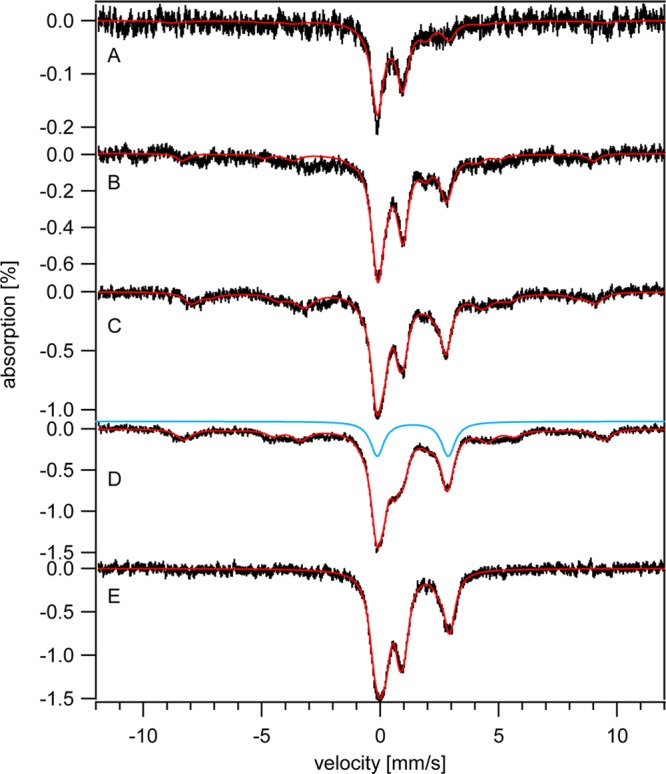
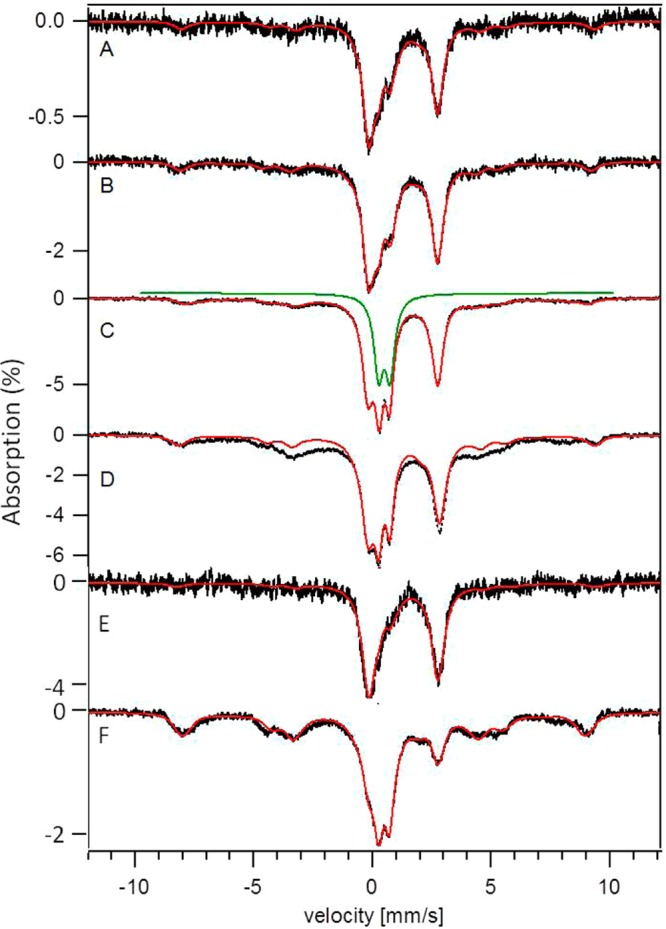
Strains lacking and overexpressing the vacuolar iron (Fe) importer CCC1 were characterized using Mössbauer and EPR spectroscopies. Vacuolar Fe import is impeded in Δccc1 cells and enhanced in CCC1-up cells, causing vacuolar Fe in these strains to decline and accumulate, respectively, relative to WT cells. Cytosolic Fe levels should behave oppositely. The Fe content of Δccc1 cells grown under low-Fe conditions was similar to that in WT cells. Most Fe was mitochondrial with some nonheme high spin (NHHS) Fe(II) present. Δccc1 cells grown with increasing Fe concentration in the medium contained less total Fe, less vacuolar HS Fe(III), and more NHHS Fe(II) than in comparable WT cells. As the Fe concentration in the growth medium increased, the concentration of HS Fe(III) in Δccc1 cells increased to just 60% of WT levels, while NHHS Fe(II) increased to twice WT levels, suggesting that the NHHS Fe(II) was cytosolic. Δccc1 cells suffered more oxidative damage than WT cells, suggesting that the accumulated NHHS Fe(II) promoted Fenton chemistry. The Fe concentration in CCC1-up cells was higher than in WT cells; the extra Fe was present as NHHS Fe(II) and Fe(III) and as Fe(III) oxyhydroxide nanoparticles. These cells contained less mitochondrial Fe and exhibited less ROS damage than Δccc1 cells. CCC1-up cells were adenine-deficient on minimal medium; supplementing with adenine caused a decline of NHHS Fe(II) suggesting that some of the NHHS Fe(II) that accumulated in these cells was associated with adenine deficiency rather than the overexpression of CCC1. A mathematical model was developed that simulated changes in Fe distributions. Simulations suggested that only a modest proportion of the observed NHHS Fe(II) in both strains was the cytosolic form of Fe that is sensed by the Fe import regulatory system. The remainder is probably generated by the reduction of the vacuolar NHHS Fe(III) species.
Figures





References
-
- Crichton R. R.; Pierre J. L. (2001) Old iron, young copper: From Mars to Venus. Biometals 14, 99–112. - PubMed
-
- Flint D. H.; Allen R. M. (1996) Iron-sulfur proteins with nonredox functions. Chem. Rev. 96, 2315–2334. - PubMed
-
- Lill R.; Hoffmann B.; Molik S.; Pierik A. J.; Rietzschel N.; Stehling O.; Uzarska M. A.; Webert H.; Wilbrecht C.; Mühlenhoff U. (2012) The role of mitochondria in cellular iron-sulfur protein biogenesis and iron metabolism. Biochim. Biophys. Acta 1823, 1491–1508. - PubMed
-
- Li L. T.; Chen O. S.; Ward D. M.; Kaplan J. (2001) CCC1 is a transporter that mediates vacuolar iron storage in yeast. J. Biol. Chem. 276, 29515–29519. - PubMed
-
- Urbanowski J. L.; Piper R. C. (1999) The iron transporter fth1p forms a complex with the Fet5 iron oxidase and resides on the vacuolar membrane. J. Biol. Chem. 274, 38061–38070. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
