

The TORC1 inhibitors Nprl2 and Nprl3 mediate an adaptive response to amino-acid starvation in Drosophila
- PMID: 24786828
- PMCID: PMC4131179
- DOI: 10.1038/cdd.2014.63
The TORC1 inhibitors Nprl2 and Nprl3 mediate an adaptive response to amino-acid starvation in Drosophila
Abstract
Target of rapamycin complex 1 (TORC1) is a master regulator of metabolism in eukaryotes that integrates information from multiple upstream signaling pathways. In yeast, the Nitrogen permease regulators 2 and 3 (Npr2 and Npr3) mediate an essential response to amino-acid limitation upstream of TORC1. In mammals, the Npr2 ortholog, Nprl2, is a putative tumor suppressor gene that inhibits cell growth and enhances sensitivity to numerous anticancer drugs including cisplatin. However, the precise role of Nprl2 and Nprl3 in the regulation of metabolism in metazoans remains poorly defined. Here we demonstrate that the central importance of Nprl2 and Nprl3 in the response to amino-acid starvation has been conserved from single celled to multicellular animals. We find that in Drosophila Nprl2 and Nprl3 physically interact and are targeted to lysosomes and autolysosomes. Using oogenesis as a model system, we show that Nprl2 and Nprl3 inhibit TORC1 signaling in the female germline in response to amino-acid starvation. Moreover, the inhibition TORC1 by Nprl2/3 is critical to the preservation of female fertility during times of protein scarcity. In young egg chambers the failure to downregulate TORC1 in response to amino-acid limitation triggers apoptosis. Thus, our data suggest the presence of a metabolic checkpoint that initiates a cell death program when TORC1 activity remains inappropriately high during periods of amino-acid and/or nutrient scarcity in oogenesis. Finally, we demonstrate that Nprl2/3 work in concert with the TORC1 inhibitors Tsc1/2 to fine tune TORC1 activity during oogenesis and that Tsc1 is a critical downstream effector of Akt1 in the female germline.
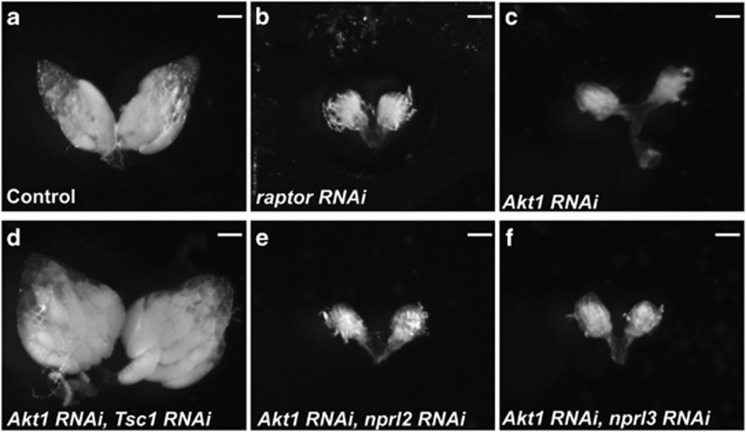
Figures





References
-
- King RC. The meiotic behavior of the Drosophila oocyte. Int Rev Cytol. 1970;28:125–168. - PubMed
-
- Drummond-Barbosa D, Spradling AC. Stem cells and their progeny respond to nutritional changes during Drosophila oogenesis. Dev Biol. 2001;231:265–278. - PubMed
-
- LaFever L, Drummond-Barbosa D. Direct control of germline stem cell division and cyst growth by neural insulin in Drosophila. Science. 2005;309:1071–1073. - PubMed
-
- Spradling AC.In: Bate M, Martinez-Arias A (eds). The Development of Drosophila melanogaster Cold Spring Harbor Laboratory Press: NY, USA; 19931–70.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
