

Interaction of heat shock protein 70 with membranes depends on the lipid environment
- PMID: 24789271
- PMCID: PMC4389847
- DOI: 10.1007/s12192-014-0511-x
Interaction of heat shock protein 70 with membranes depends on the lipid environment
Abstract
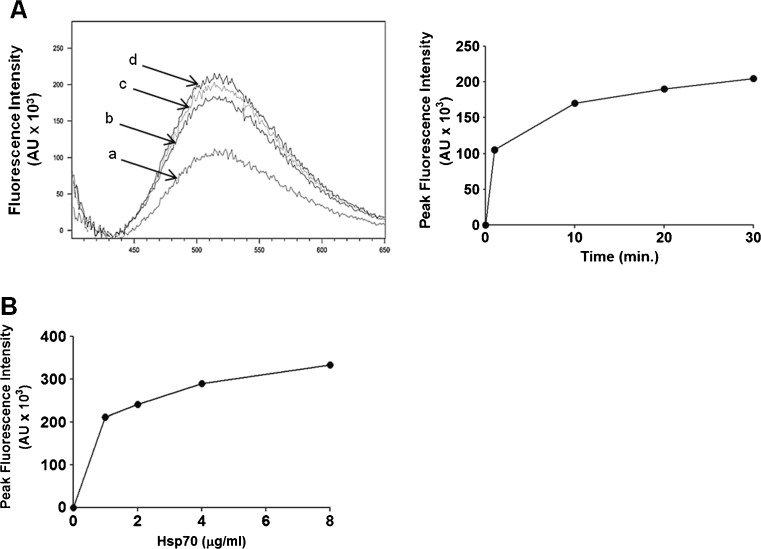
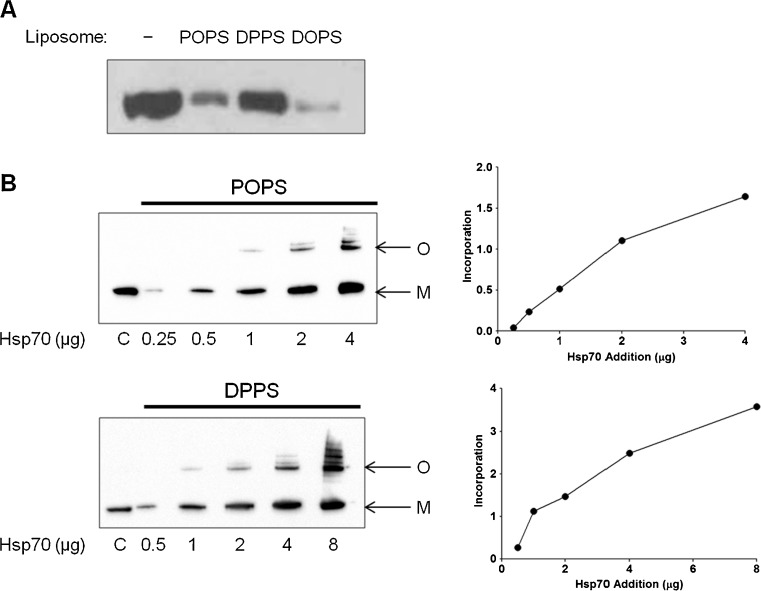
Heat shock proteins (hsp) are well recognized for their protein folding activity. Additionally, hsp expression is enhanced during stress conditions to preserve cellular homeostasis. Hsp are also detected outside cells, released by an active mechanism independent of cell death. Extracellular hsp appear to act as signaling molecules as part of a systemic response to stress. Extracellular hsp do not contain a consensus signal for their secretion via the classical ER-Golgi compartment. Therefore, they are likely exported by an alternative mechanism requiring translocation across the plasma membrane. Since Hsp70, the major inducible hsp, has been detected on surface of stressed cells, we propose that membrane interaction is the first step in the export process. The question that emerges is how does this charged cytosolic protein interact with lipid membranes? Prior studies have shown that Hsp70 formed ion conductance pathways within artificial lipid bilayers. These early observations have been extended herewith using a liposome insertion assay. We showed that Hsp70 selectively interacted with negatively charged phospholipids, particularly phosphatidyl serine (PS), within liposomes, which was followed by insertion into the lipid bilayer, forming high-molecular weight oligomers. Hsp70 displayed a preference for less fluid lipid environments and the region embedded into the lipid membrane was mapped toward the C-terminus end of the molecule. The results from our studies provide evidence of an unexpected ability of a large, charged protein to become inserted into a lipid membrane. This observation provides a new paradigm for the interaction of proteins with lipid environments. In addition, it may explain the export mechanism of an increasing number of proteins that lack the consensus secretory signals.
Figures


References
-
- Aprile FA, Dhulesia A, Stengel F, Roodveldt C, Benesch JL, Tortora P, Robinson CV, Salvatella X, Dobson CM, Cremades N. Hsp70 oligomerization is mediated by an interaction between the interdomain linker and the substrate-binding domain. PLoS One. 2013;8:e67961. doi: 10.1371/journal.pone.0067961. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
