

Bioenergetic constraints on the evolution of complex life
- PMID: 24789818
- PMCID: PMC3996473
- DOI: 10.1101/cshperspect.a015982
Bioenergetic constraints on the evolution of complex life
Abstract
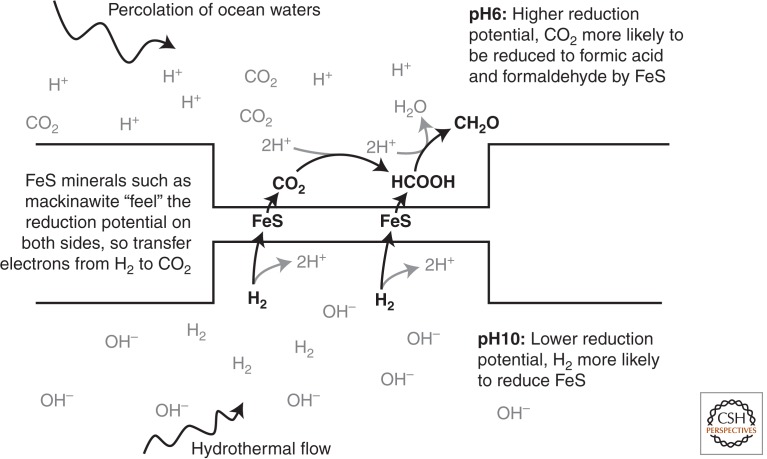
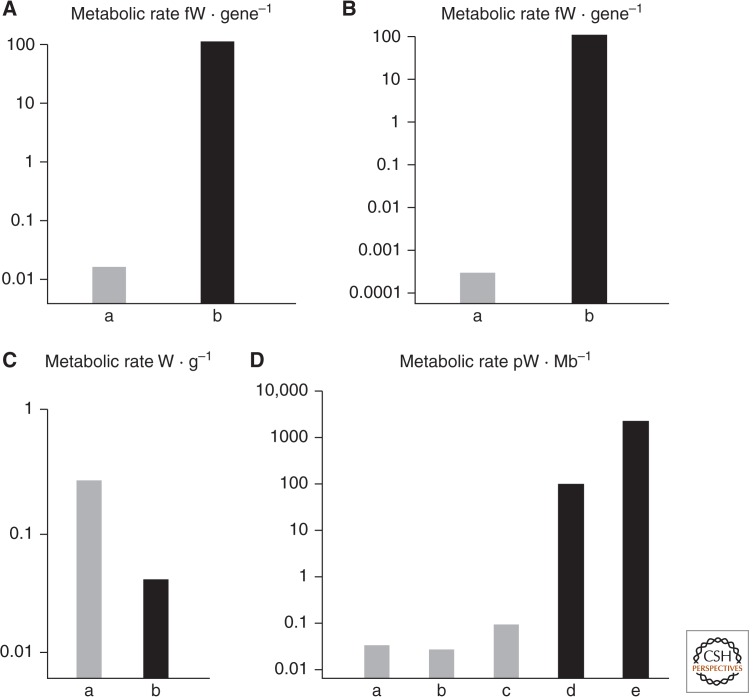
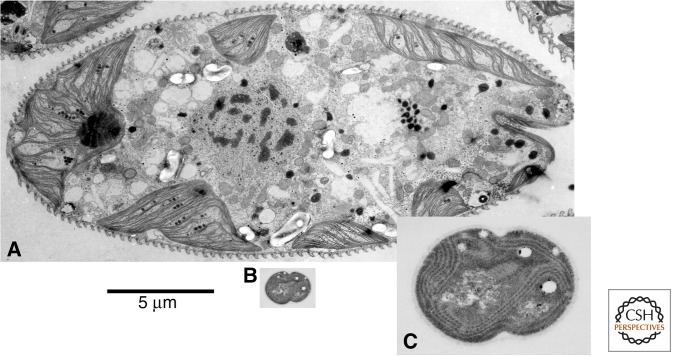
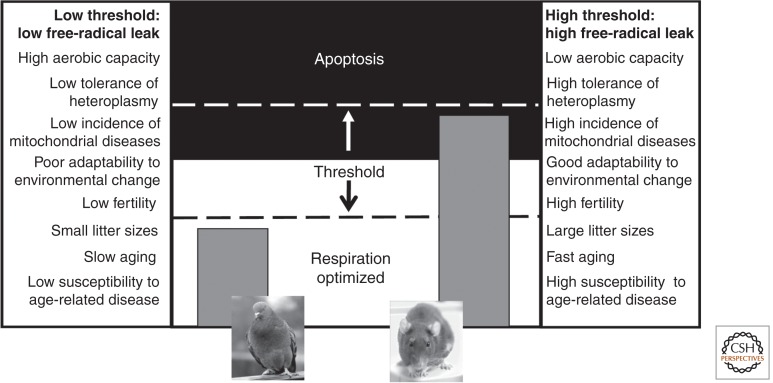
All morphologically complex life on Earth, beyond the level of cyanobacteria, is eukaryotic. All eukaryotes share a common ancestor that was already a complex cell. Despite their biochemical virtuosity, prokaryotes show little tendency to evolve eukaryotic traits or large genomes. Here I argue that prokaryotes are constrained by their membrane bioenergetics, for fundamental reasons relating to the origin of life. Eukaryotes arose in a rare endosymbiosis between two prokaryotes, which broke the energetic constraints on prokaryotes and gave rise to mitochondria. Loss of almost all mitochondrial genes produced an extreme genomic asymmetry, in which tiny mitochondrial genomes support, energetically, a massive nuclear genome, giving eukaryotes three to five orders of magnitude more energy per gene than prokaryotes. The requirement for endosymbiosis radically altered selection on eukaryotes, potentially explaining the evolution of unique traits, including the nucleus, sex, two sexes, speciation, and aging.
Figures
References
-
- Allen JF 1993. Control of gene expression by redox potential and the requirement for chloroplast and mitochondrial genomes. J Theor Biol 165: 609–631 - PubMed
-
- Amend JP, McCollom TM 2009. Energetics of biomolecule synthesis on early Earth. In Chemical evolution II: From the origins of life to modern society (ed. Zaikowski L, et al. ), pp. 63–94 American Chemical Society, Washington, DC
-
- Andersson SG, Kurland CG 1999. Origins of mitochondria and hydrogenosomes. Curr Opin Microbiol 2: 535–541 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources