

Distinct assembly profiles of HLA-B molecules
- PMID: 24790147
- PMCID: PMC4117407
- DOI: 10.4049/jimmunol.1301670
Distinct assembly profiles of HLA-B molecules
Abstract
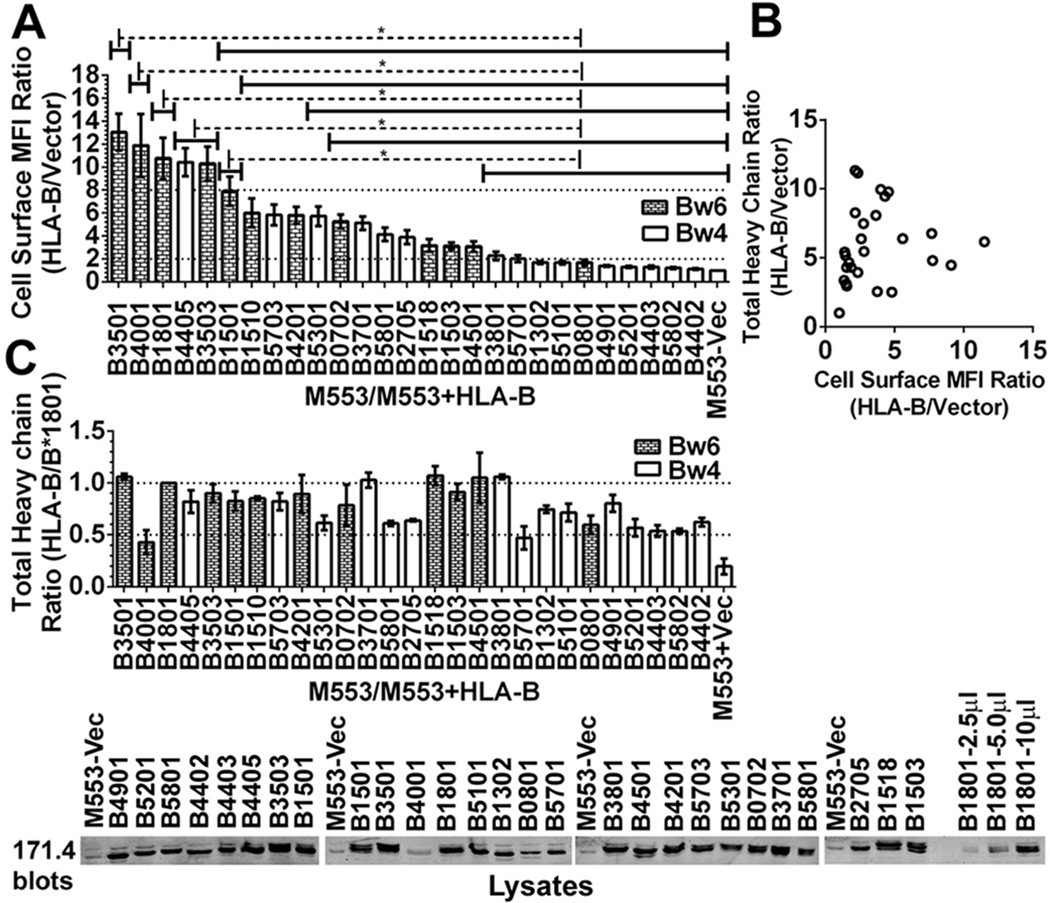
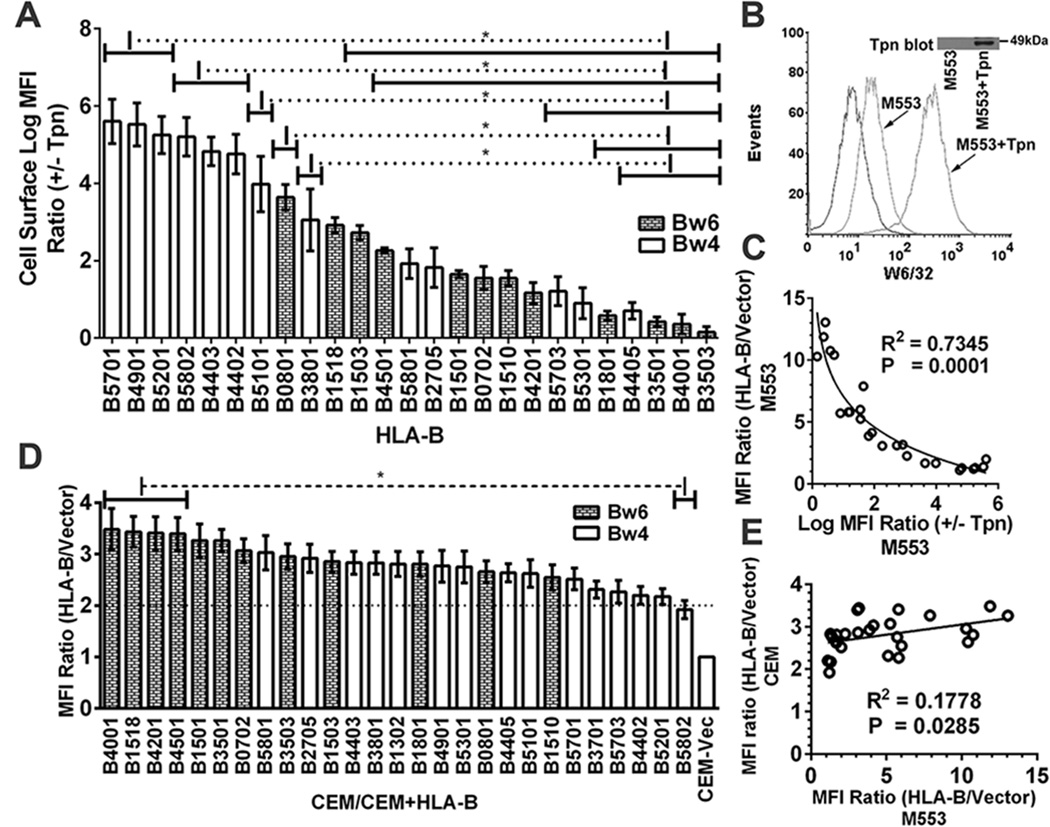
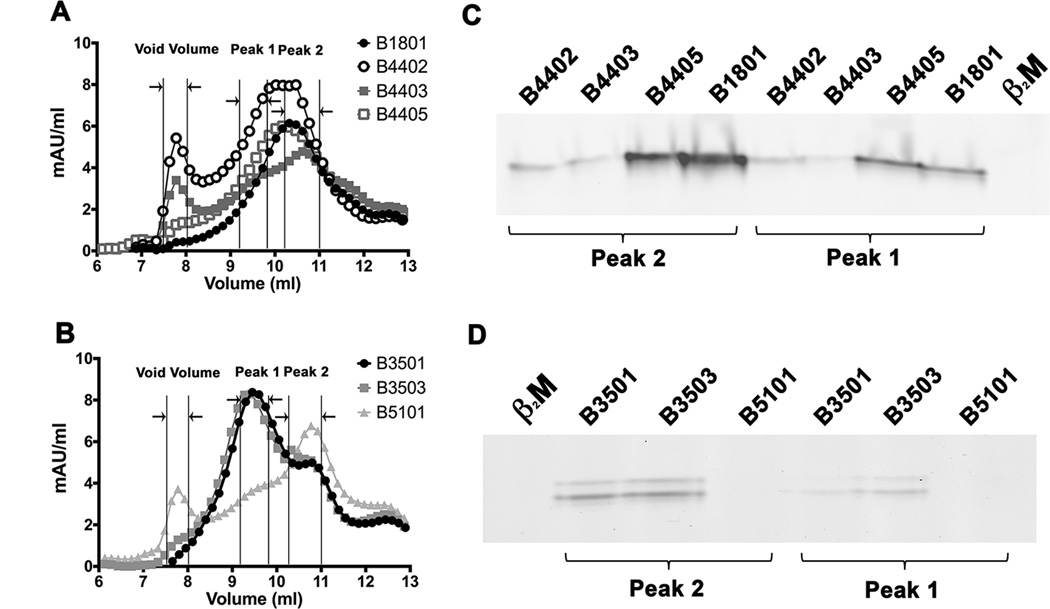
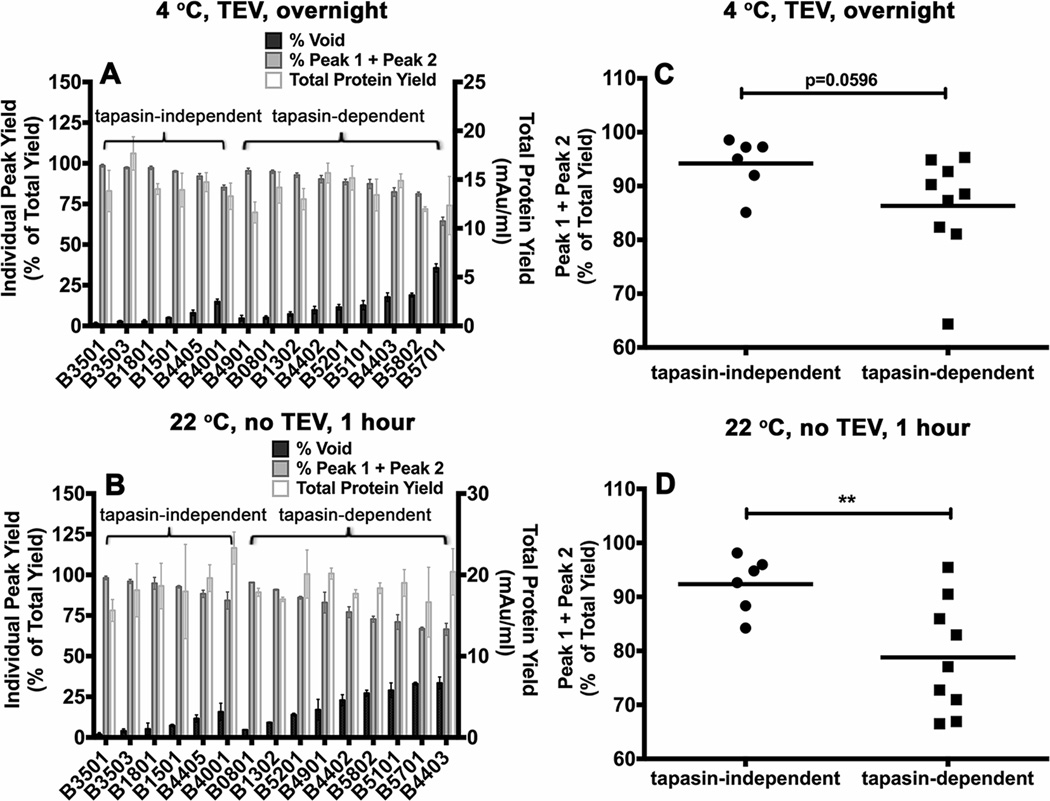
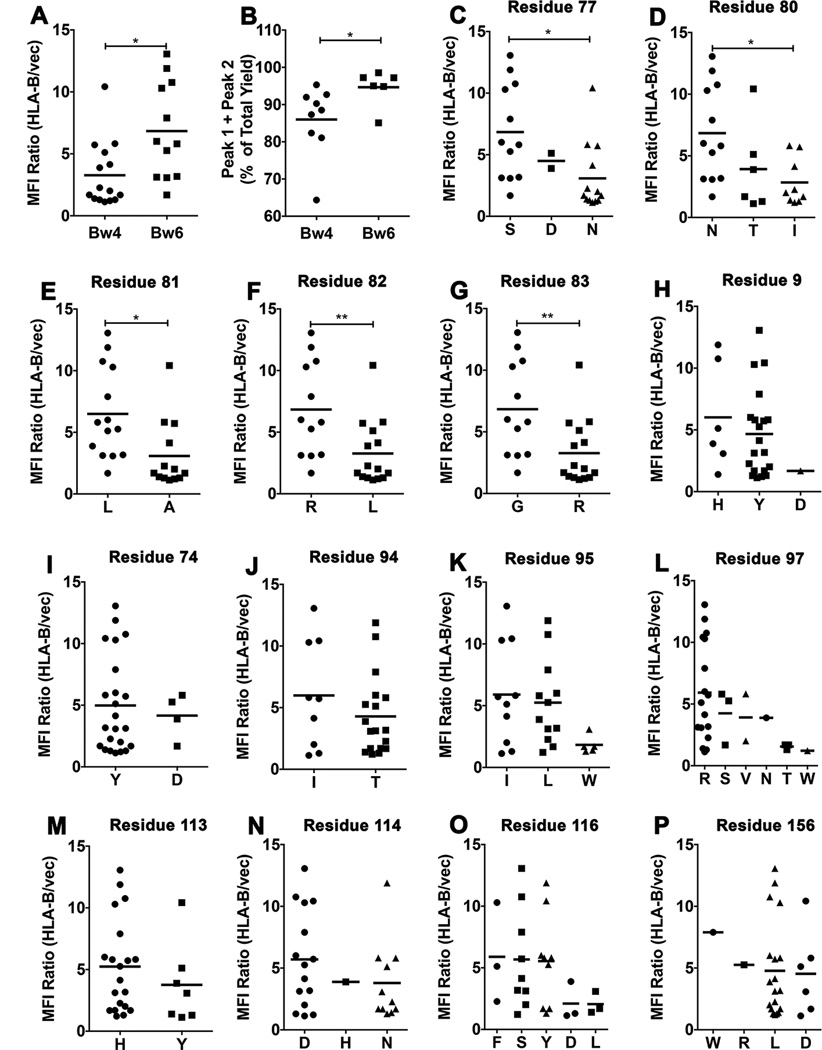
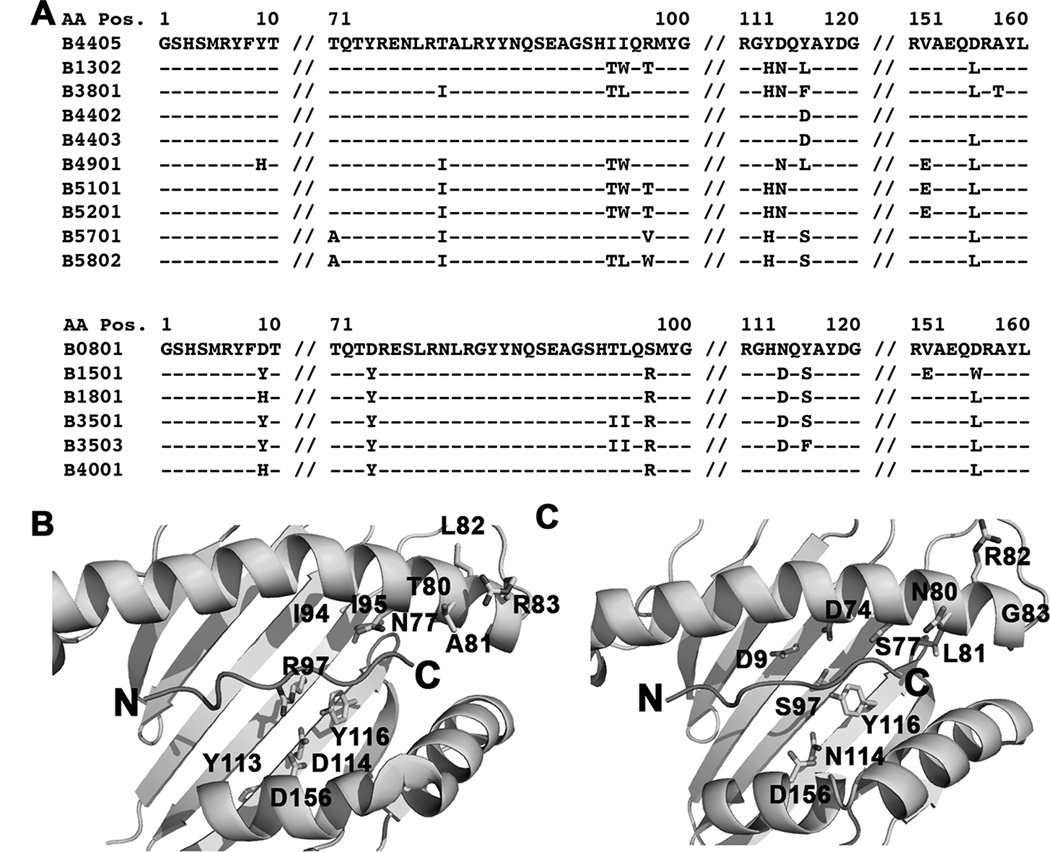
MHC class I polymorphisms are known to influence outcomes in a number of infectious diseases, cancers, and inflammatory diseases. Human MHC class I H chains are encoded by the HLA-A, HLA-B, and HLA-C genes. These genes are highly polymorphic, with the HLA-B locus being the most variable. Each HLA class I protein binds to a distinct set of peptide Ags, which are presented to CD8(+) T cells. HLA-disease associations have been shown in some cases to link to the peptide-binding characteristics of individual HLA class I molecules. In this study, we show that polymorphisms at the HLA-B locus profoundly influence the assembly characteristics of HLA-B molecules and the stabilities of their peptide-deficient forms. In particular, dependence on the assembly factor tapasin is highly variable, with frequent occurrence of strongly tapasin-dependent or independent allotypes. Several polymorphic HLA-B residues located near the C-terminal end of the peptide are key determinants of tapasin-independent assembly. In vitro refolded forms of tapasin-independent allotypes assemble more readily with peptides compared to tapasin-dependent allotypes that belong to the same supertype, and, during refolding, reduced aggregation of tapasin-independent allotypes is observed. Paradoxically, in HIV-infected individuals, greater tapasin-independent HLA-B assembly confers more rapid progression to death, consistent with previous findings that some HLA-B allotypes shown to be tapasin independent are associated with rapid progression to multiple AIDS outcomes. Together, these findings demonstrate significant variations in the assembly of HLA-B molecules and indicate influences of HLA-B-folding patterns upon infectious disease outcomes.
Figures
Similar articles
-
HLA-B polymorphisms and intracellular assembly modes.Mol Immunol. 2015 Dec;68(2 Pt A):89-93. doi: 10.1016/j.molimm.2015.07.007. Epub 2015 Jul 31. Mol Immunol. 2015. PMID: 26239417 Free PMC article. Review.
-
Distinct functions of tapasin revealed by polymorphism in MHC class I peptide loading.J Immunol. 2000 Jan 1;164(1):292-9. doi: 10.4049/jimmunol.164.1.292. J Immunol. 2000. PMID: 10605023
-
HLA tapasin independence: broader peptide repertoire and HIV control.Proc Natl Acad Sci U S A. 2020 Nov 10;117(45):28232-28238. doi: 10.1073/pnas.2013554117. Epub 2020 Oct 23. Proc Natl Acad Sci U S A. 2020. PMID: 33097667 Free PMC article.
-
Assembly and intracellular trafficking of HLA-B*3501 and HLA-B*3503.Immunogenetics. 2009 Dec;61(11-12):703-16. doi: 10.1007/s00251-009-0399-2. Immunogenetics. 2009. PMID: 19838694 Free PMC article.
-
Polymorphisms of HLA-B: influences on assembly and immunity.Curr Opin Immunol. 2020 Jun;64:137-145. doi: 10.1016/j.coi.2020.05.008. Epub 2020 Jun 30. Curr Opin Immunol. 2020. PMID: 32619904 Free PMC article. Review.
Cited by
-
Relationship of HLA-B alleles on susceptibility to and protection from HIV infection in Turkish population.North Clin Istanb. 2022 Dec 13;10(1):67-73. doi: 10.14744/nci.2021.00018. eCollection 2023. North Clin Istanb. 2022. PMID: 36910436 Free PMC article.
-
Distinct Polymorphisms in HLA Class I Molecules Govern Their Susceptibility to Peptide Editing by TAPBPR.Cell Rep. 2019 Nov 5;29(6):1621-1632.e3. doi: 10.1016/j.celrep.2019.09.074. Cell Rep. 2019. PMID: 31693900 Free PMC article.
-
Exchange catalysis by tapasin exploits conserved and allele-specific features of MHC-I molecules.Nat Commun. 2021 Jul 9;12(1):4236. doi: 10.1038/s41467-021-24401-4. Nat Commun. 2021. PMID: 34244493 Free PMC article.
-
TAPBPR bridges UDP-glucose:glycoprotein glucosyltransferase 1 onto MHC class I to provide quality control in the antigen presentation pathway.Elife. 2017 Apr 20;6:e23049. doi: 10.7554/eLife.23049. Elife. 2017. PMID: 28425917 Free PMC article.
-
Selector function of MHC I molecules is determined by protein plasticity.Sci Rep. 2015 Oct 20;5:14928. doi: 10.1038/srep14928. Sci Rep. 2015. PMID: 26482009 Free PMC article.
References
-
- Neefjes J, Jongsma ML, Paul P, Bakke O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat Rev Immunol. 2011;11:823–836. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI044115/AI/NIAID NIH HHS/United States
- U01-AI35040/AI/NIAID NIH HHS/United States
- U01-AI35042/AI/NIAID NIH HHS/United States
- U01 AI035041/AI/NIAID NIH HHS/United States
- UM1 AI035043/AI/NIAID NIH HHS/United States
- AI044115/AI/NIAID NIH HHS/United States
- U01-AI35041/AI/NIAID NIH HHS/United States
- U01-AI35039/AI/NIAID NIH HHS/United States
- U01 AI035040/AI/NIAID NIH HHS/United States
- U01 AI035039/AI/NIAID NIH HHS/United States
- UM1-AI35043/AI/NIAID NIH HHS/United States
- K07 CA140360/CA/NCI NIH HHS/United States
- U01 AI035042/AI/NIAID NIH HHS/United States
- R56 AI044115/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
