

All-trans retinoic acid induces arginase-1 and inducible nitric oxide synthase-producing dendritic cells with T cell inhibitory function
- PMID: 24790153
- PMCID: PMC4032093
- DOI: 10.4049/jimmunol.1303073
All-trans retinoic acid induces arginase-1 and inducible nitric oxide synthase-producing dendritic cells with T cell inhibitory function
Abstract
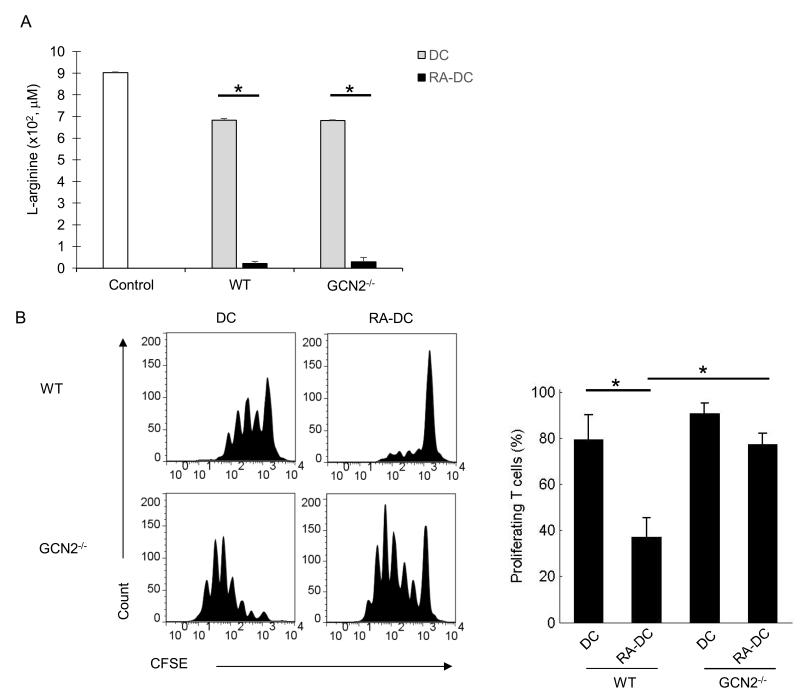
Hepatic stellate cells (HSC) are a major source of the immunoregulatory metabolite all-trans retinoic acid (ATRA), which may contribute to the generation of tolerogenic dendritic cells (DCs) in the liver. The present study seeks to clarify the mechanism(s) through which ATRA promotes the development of tolerogenic DCs. Although bone marrow-derived ATRA-treated DCs (RA-DCs) and conventional DCs had comparable surface phenotype, RA-DCs had diminished stimulatory capacity and could directly inhibit the expansion of DC/OVA-stimulated OT-II T cells. Arginase-1 (Arg-1) was found promote suppression because 1) ATRA was a potent inducer of Arg-1 protein and activity, 2) the Arg-1 inhibitor N(w)-hydroxy nor-l-arginine partially reversed suppression, and 3) the suppressive function of RA-DCs was partially compromised using OT-II T cells from GCN2(-/-) mice, which are insensitive to Arg-1. Inducible NO synthase (iNOS), however, was found to be a more significant contributor to RA-DC function because 1) ATRA potentiated the expression of IFN-γ-induced iNOS, 2) suppressive function in RA-DCs was blocked by the iNOS inhibitor N(G)-monomethyl-l-arginine, monoacetate salt, and 3) RA-DCs derived from iNOS(-/-) mice exhibited near complete loss of tolerogenic function, despite sustained Arg-1 activity. The expression of iNOS and the suppressive function of RA-DCs were dependent on both IFN-γ and ATRA. Furthermore, the in vivo behavior of RA-DCs proved to be consistent with their in vitro behavior. Thus, we conclude that ATRA enhances both Arg-1 and iNOS expression in IFN-γ-treated DCs, resulting in a tolerogenic phenotype. These findings elucidate mechanisms through which ATRA may contribute to liver immune tolerance.
Copyright © 2014 by The American Association of Immunologists, Inc.
Figures







References
-
- Yu M-C, Chen C-H, Liang X, Wang L, Gandhi CR, Fung JJ, Lu L, Qian S. Inhibition of T-cell responses by hepatic stellate cells via B7-H1-mediated T-cell apoptosis in mice. Hepatology. 2004;40:1312–1321. - PubMed
-
- Chen C-H, Kuo L-M, Chang Y, Wu W, Goldbach C, Ross MA, Stolz DB, Chen L, Fung JJ, Lu L, Qian S. In vivo immune modulatory activity of hepatic stellate cells in mice. Hepatology. 2006;44:1171–1181. - PubMed
-
- Blomhoff R, Wake K. Perisinusoidal stellate cells of the liver: important roles in retinol metabolism and fibrosis. FASEB J. 1991;5:271–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
