

Kainate receptors mediate signaling in both transient and sustained OFF bipolar cell pathways in mouse retina
- PMID: 24790183
- PMCID: PMC4004803
- DOI: 10.1523/JNEUROSCI.4941-13.2014
Kainate receptors mediate signaling in both transient and sustained OFF bipolar cell pathways in mouse retina
Abstract
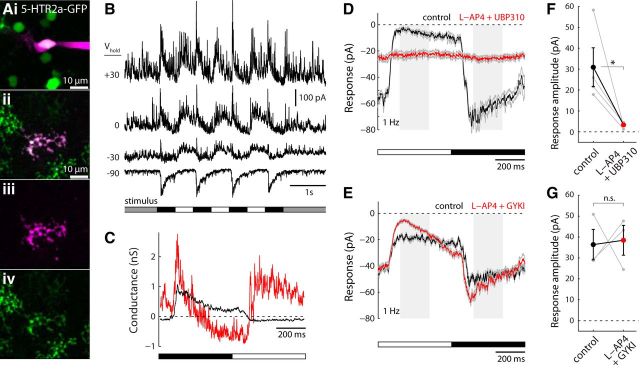
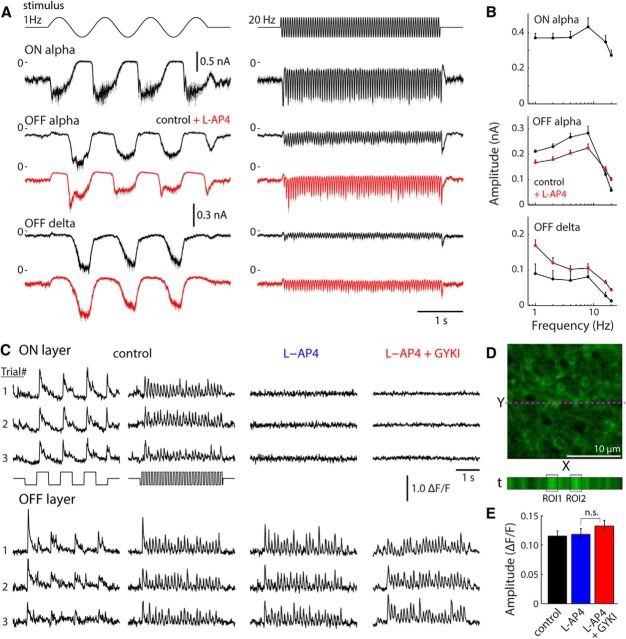
A fundamental question in sensory neuroscience is how parallel processing is implemented at the level of molecular and circuit mechanisms. In the retina, it has been proposed that distinct OFF cone bipolar cell types generate fast/transient and slow/sustained pathways by the differential expression of AMPA- and kainate-type glutamate receptors, respectively. However, the functional significance of these receptors in the intact circuit during light stimulation remains unclear. Here, we measured glutamate release from mouse bipolar cells by two-photon imaging of a glutamate sensor (iGluSnFR) expressed on postsynaptic amacrine and ganglion cell dendrites. In both transient and sustained OFF layers, cone-driven glutamate release from bipolar cells was blocked by antagonists to kainate receptors but not AMPA receptors. Electrophysiological recordings from bipolar and ganglion cells confirmed the essential role of kainate receptors for signaling in both transient and sustained OFF pathways. Kainate receptors mediated responses to contrast modulation up to 20 Hz. Light-evoked responses in all mouse OFF bipolar pathways depend on kainate, not AMPA, receptors.
Keywords: OFF bipolar cell; glutamate sensor; kainate receptor; mouse; retinal circuitry; two-photon imaging.
Conflict of interest statement
Conflict of interest: B.G.B. owns Borghuis Instruments, which sells the specialized syringe that was used for intravitreal virus injections in this study.
Figures
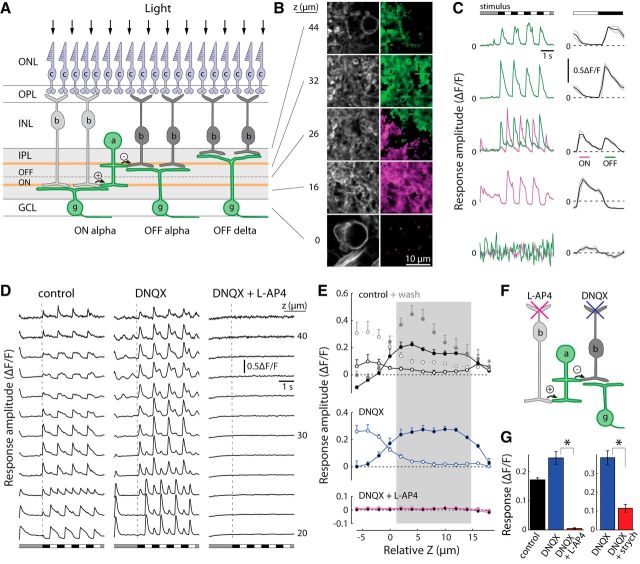
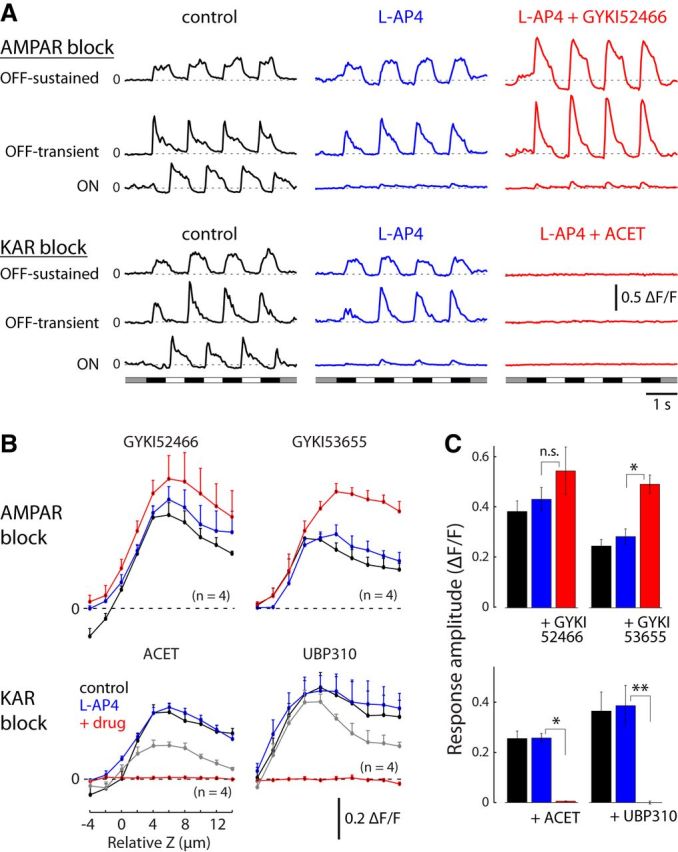
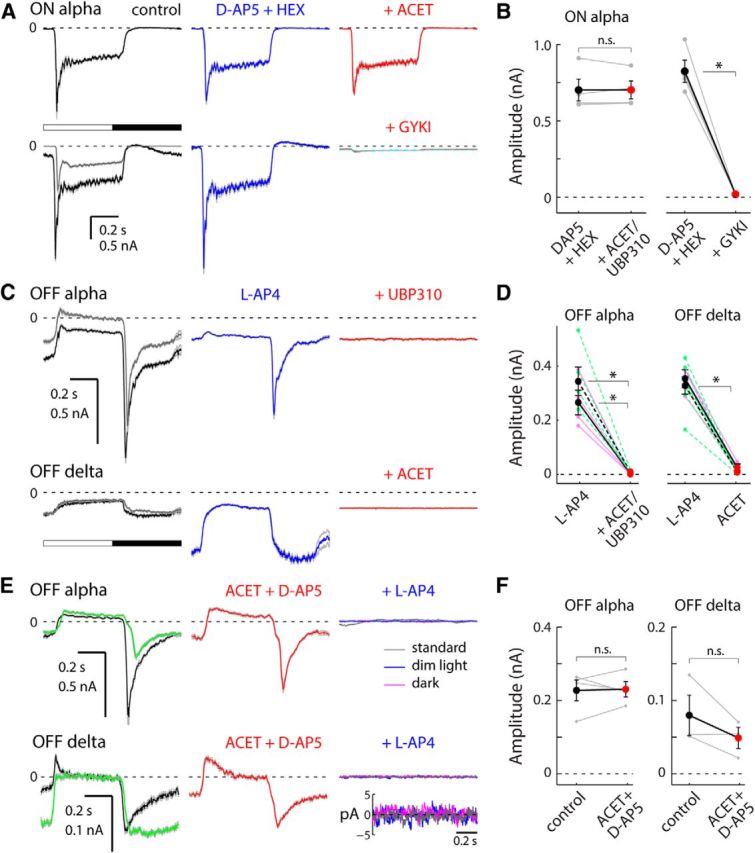
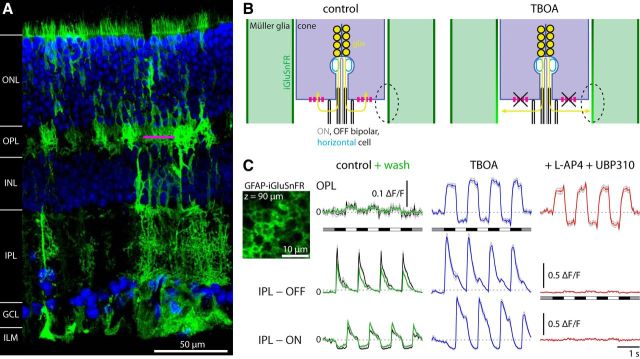
References
-
- Applebury ML, Antoch MP, Baxter LC, Chun LL, Falk JD, Farhangfar F, Kage K, Krzystolik MG, Lyass LA, Robbins JT. The murine cone photoreceptor: a single cone type expresses both S and M opsins with retinal spatial patterning. Neuron. 2000;27:513–523. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources