

Hyaluronan deficiency due to Has3 knock-out causes altered neuronal activity and seizures via reduction in brain extracellular space
- PMID: 24790187
- PMCID: PMC4004806
- DOI: 10.1523/JNEUROSCI.3458-13.2014
Hyaluronan deficiency due to Has3 knock-out causes altered neuronal activity and seizures via reduction in brain extracellular space
Abstract
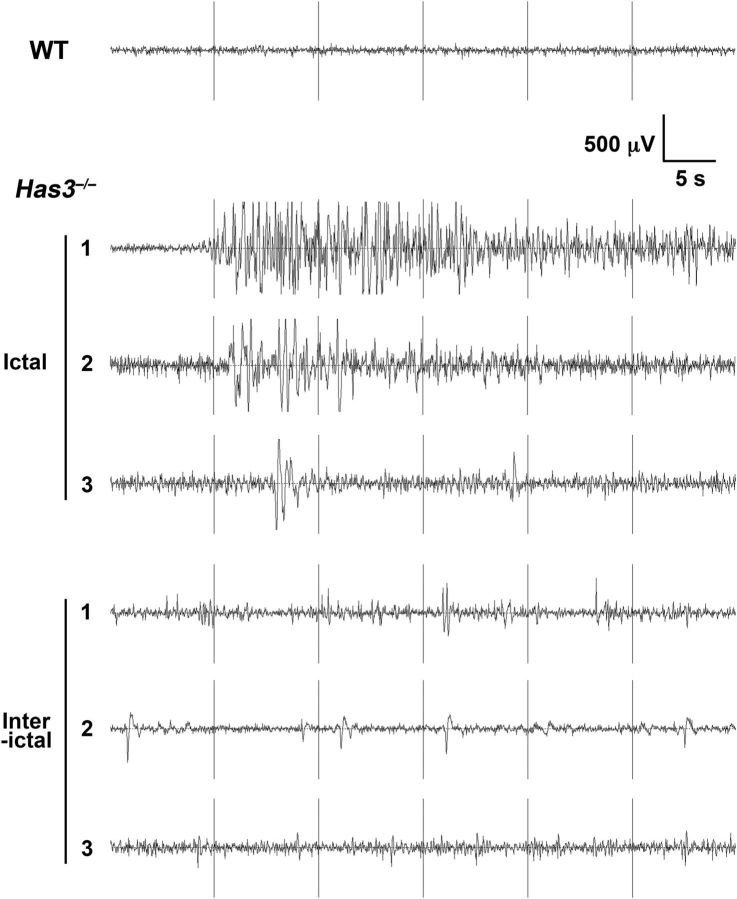
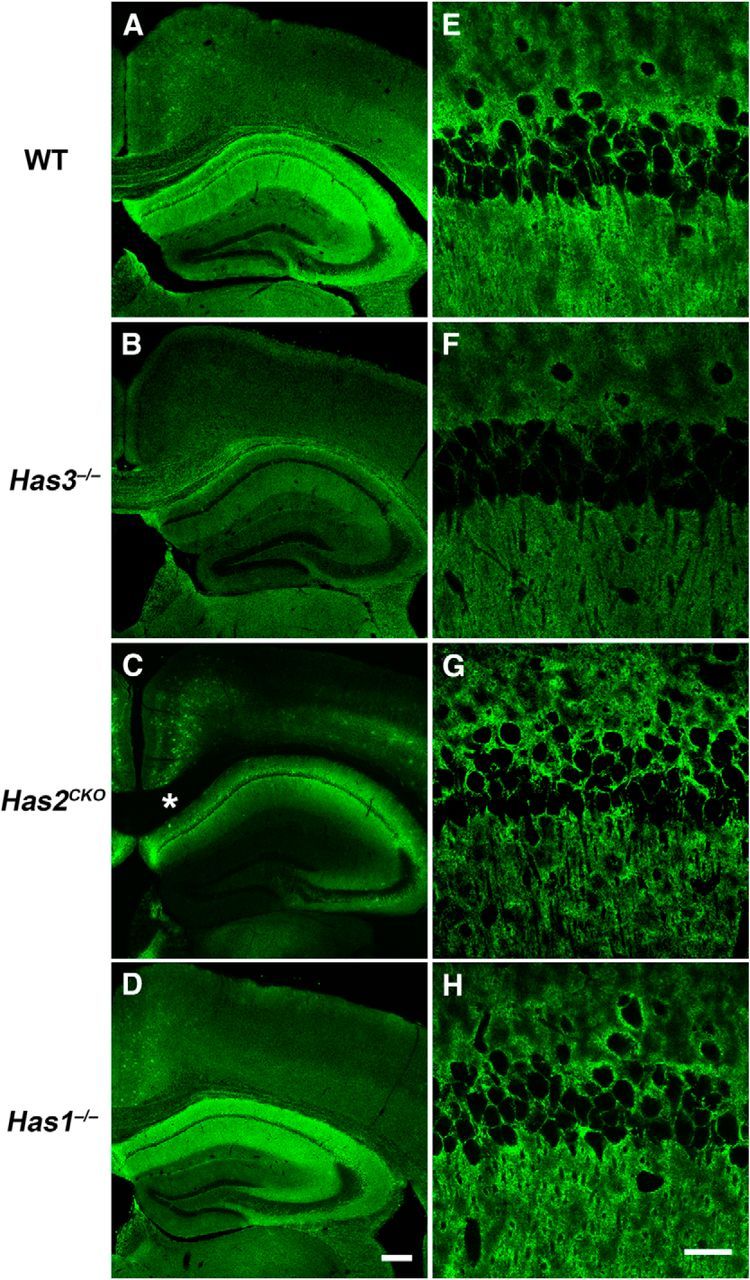
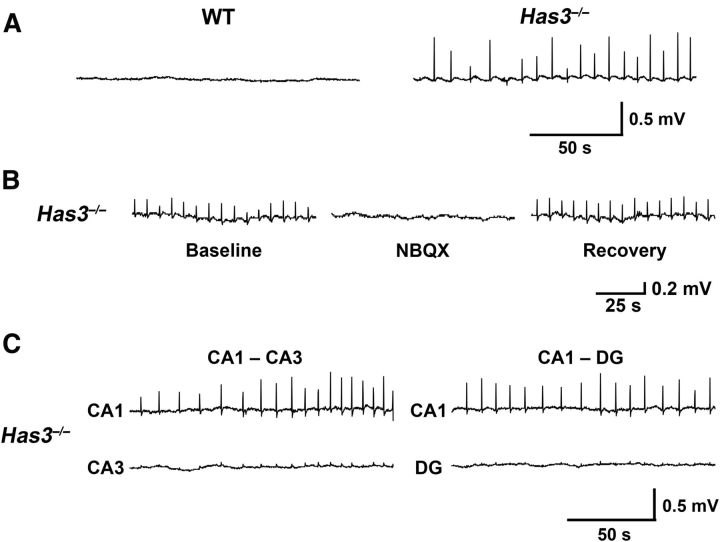
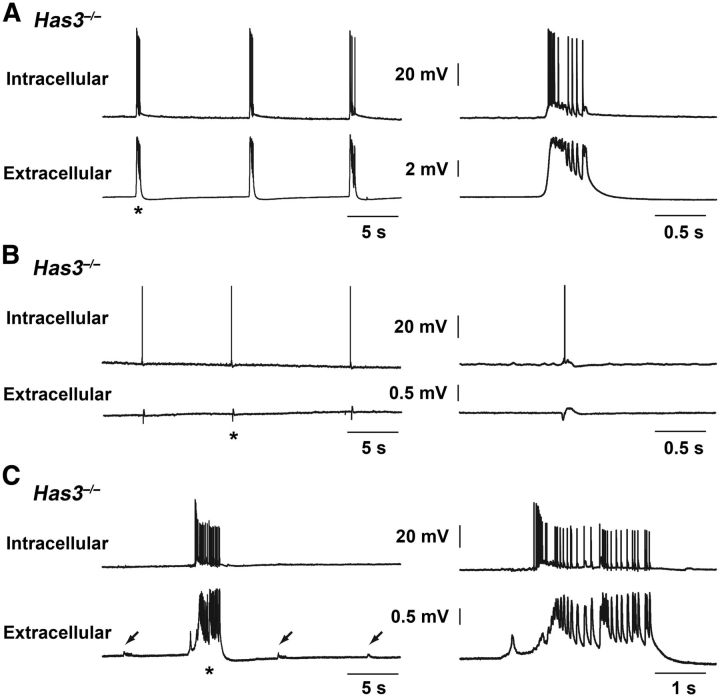
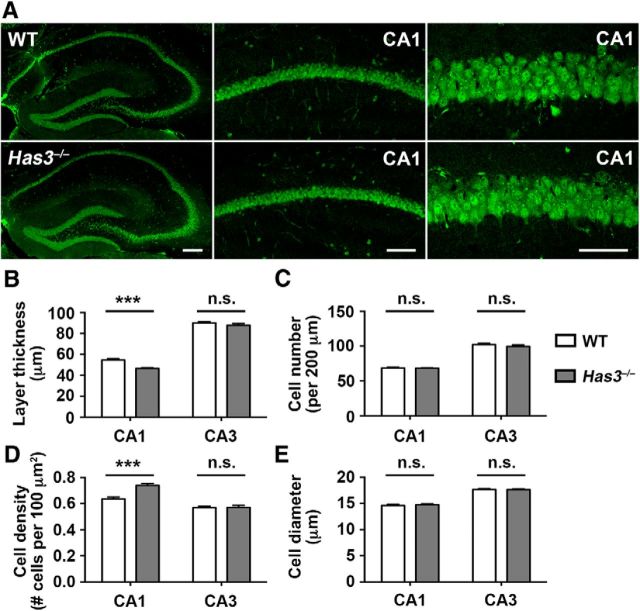
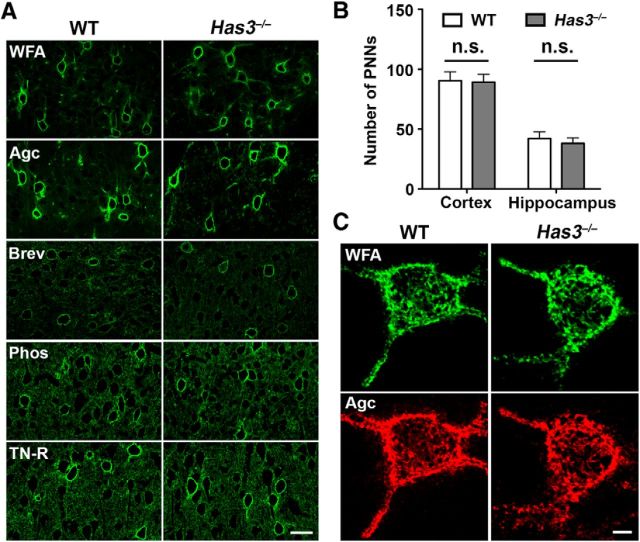
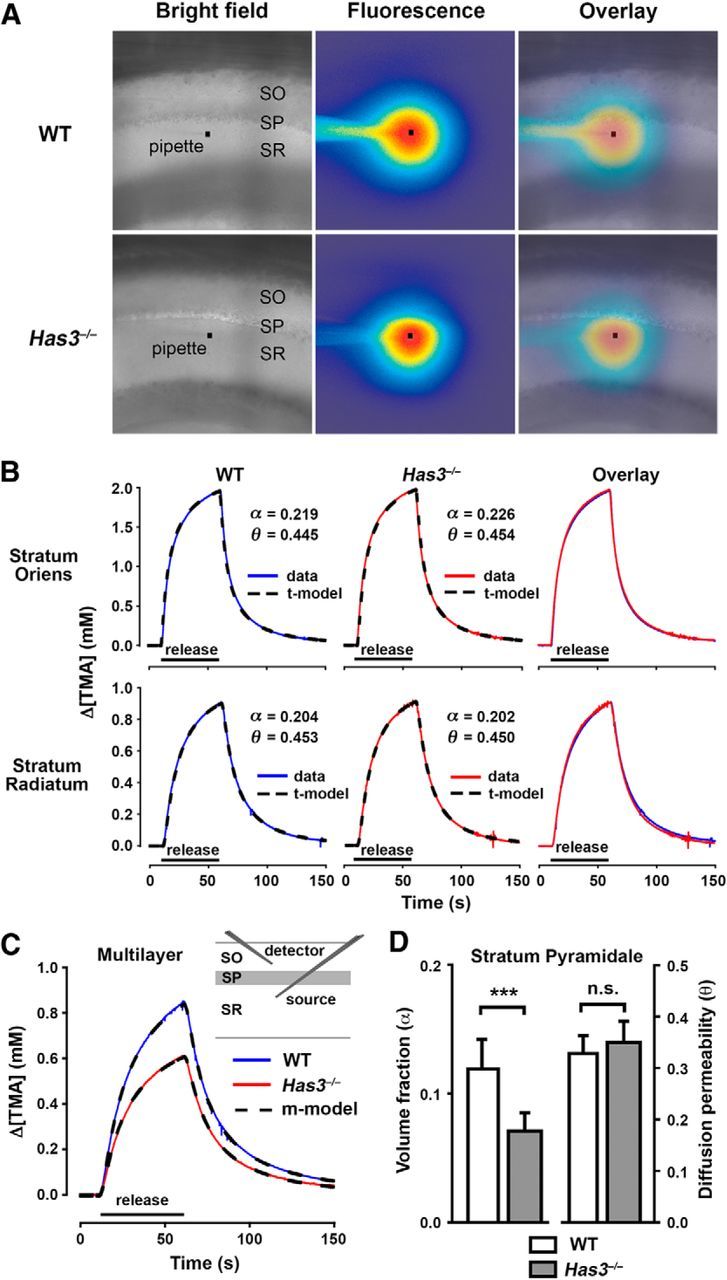
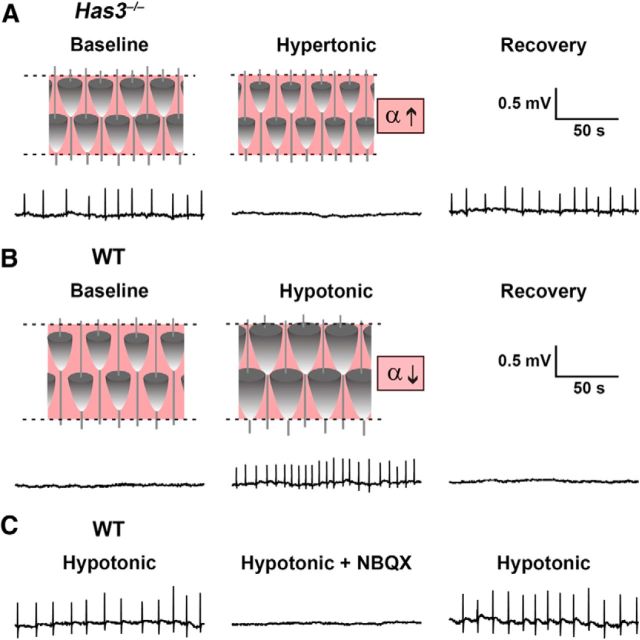
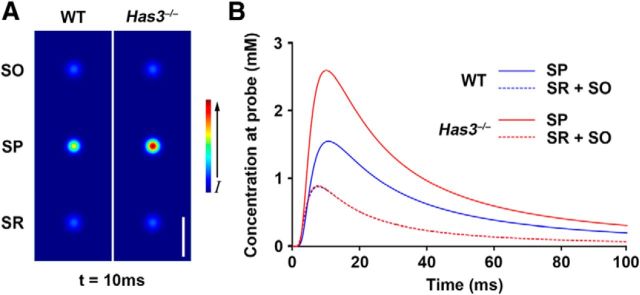
Hyaluronan (HA), a large anionic polysaccharide (glycosaminoglycan), is a major constituent of the extracellular matrix of the adult brain. To address its function, we examined the neurophysiology of knock-out mice deficient in hyaluronan synthase (Has) genes. Here we report that these Has mutant mice are prone to epileptic seizures, and that in Has3(-/-) mice, this phenotype is likely derived from a reduction in the size of the brain extracellular space (ECS). Among the three Has knock-out models, namely Has3(-/-), Has1(-/-), and Has2(CKO), the seizures were most prevalent in Has3(-/-) mice, which also showed the greatest HA reduction in the hippocampus. Electrophysiology in Has3(-/-) brain slices demonstrated spontaneous epileptiform activity in CA1 pyramidal neurons, while histological analysis revealed an increase in cell packing in the CA1 stratum pyramidale. Imaging of the diffusion of a fluorescent marker revealed that the transit of molecules through the ECS of this layer was reduced. Quantitative analysis of ECS by the real-time iontophoretic method demonstrated that ECS volume was selectively reduced in the stratum pyramidale by ∼ 40% in Has3(-/-) mice. Finally, osmotic manipulation experiments in brain slices from Has3(-/-) and wild-type mice provided evidence for a causal link between ECS volume and epileptiform activity. Our results provide the first direct evidence for the physiological role of HA in the regulation of ECS volume, and suggest that HA-based preservation of ECS volume may offer a novel avenue for development of antiepileptogenic treatments.
Figures
References
-
- Baraban SC, Southwell DG, Estrada RC, Jones DL, Sebe JY, Alfaro-Cervello C, García-Verdugo JM, Rubenstein JL, Alvarez-Buylla A. Reduction of seizures by transplantation of cortical GABAergic interneuron precursors into Kv1.1 mutant mice. Proc Natl Acad Sci U S A. 2009;106:15472–15477. doi: 10.1073/pnas.0900141106. - DOI - PMC - PubMed
-
- Bates EJ, Harper GS, Lowther DA, Preston BN. Effect of oxygen-derived reactive species on cartilage proteoglycan-hyaluronate aggregates. Biochem Int. 1984;8:629–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous