

Impaired path integration and grid cell spatial periodicity in mice lacking GluA1-containing AMPA receptors
- PMID: 24790195
- PMCID: PMC6608102
- DOI: 10.1523/JNEUROSCI.4330-13.2014
Impaired path integration and grid cell spatial periodicity in mice lacking GluA1-containing AMPA receptors
Abstract
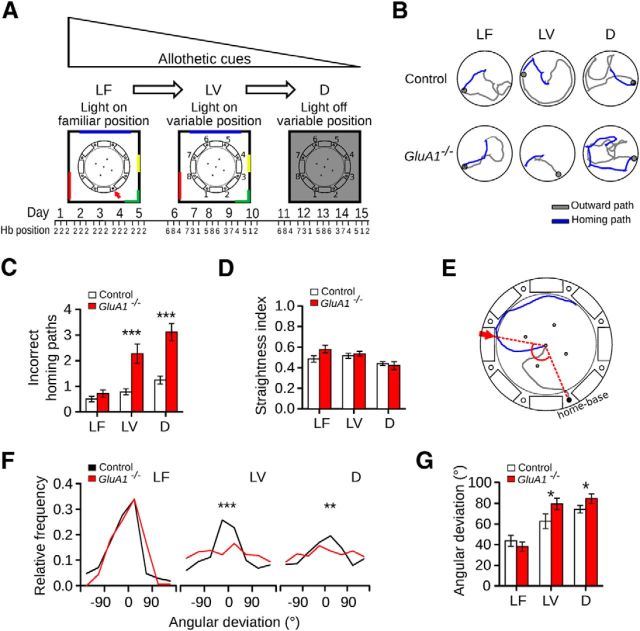
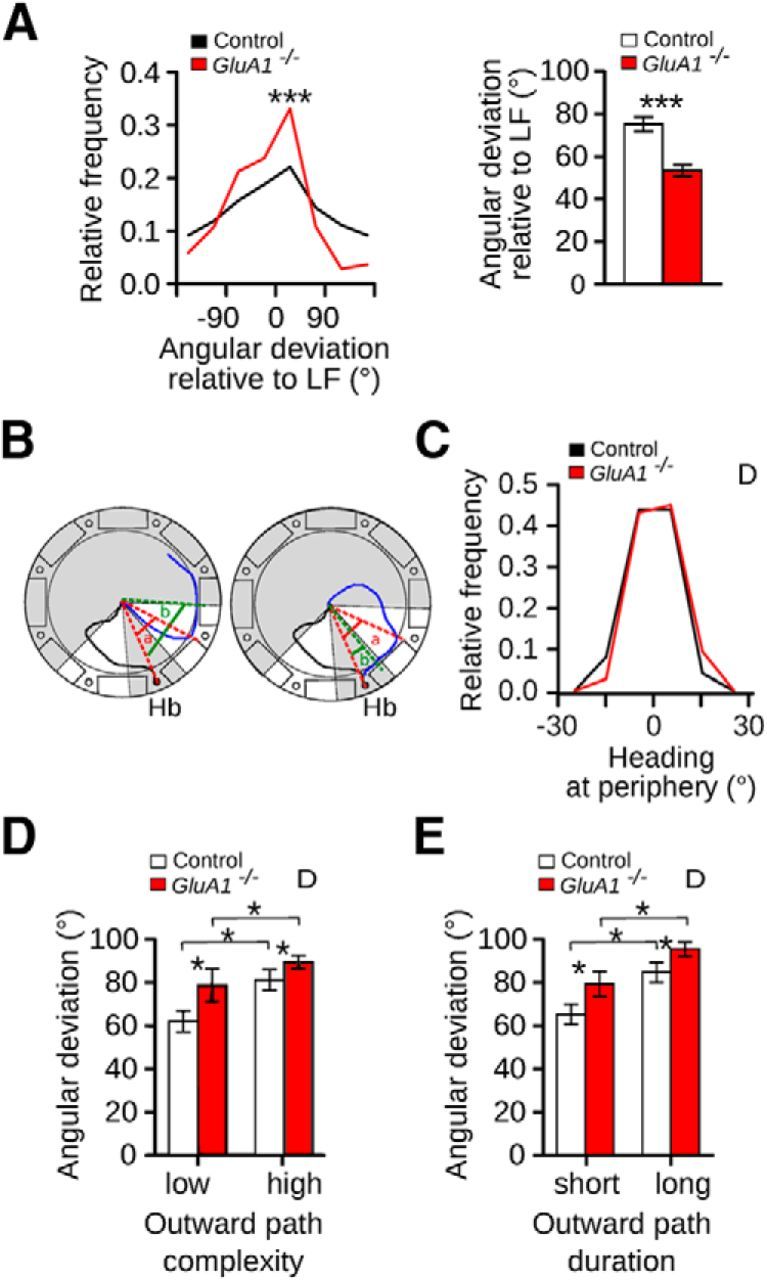
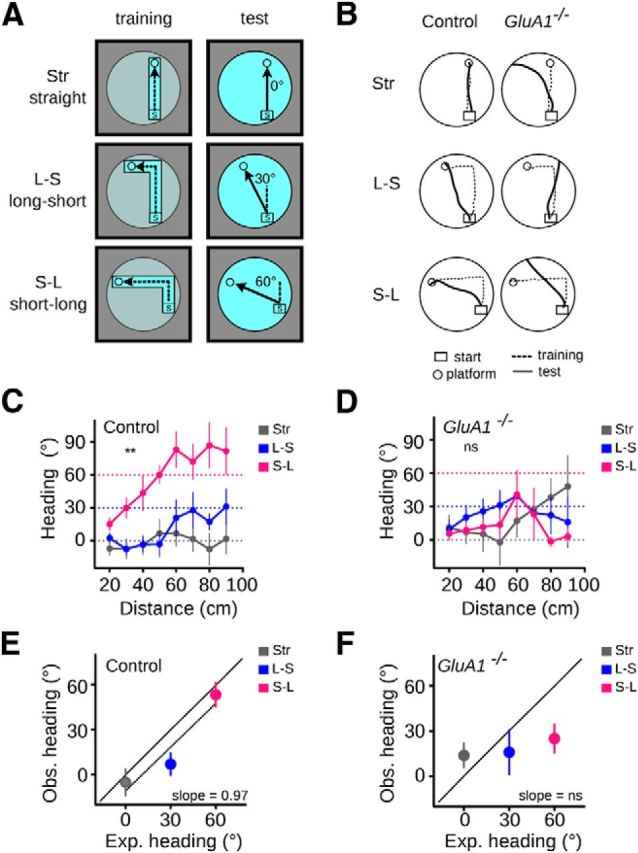
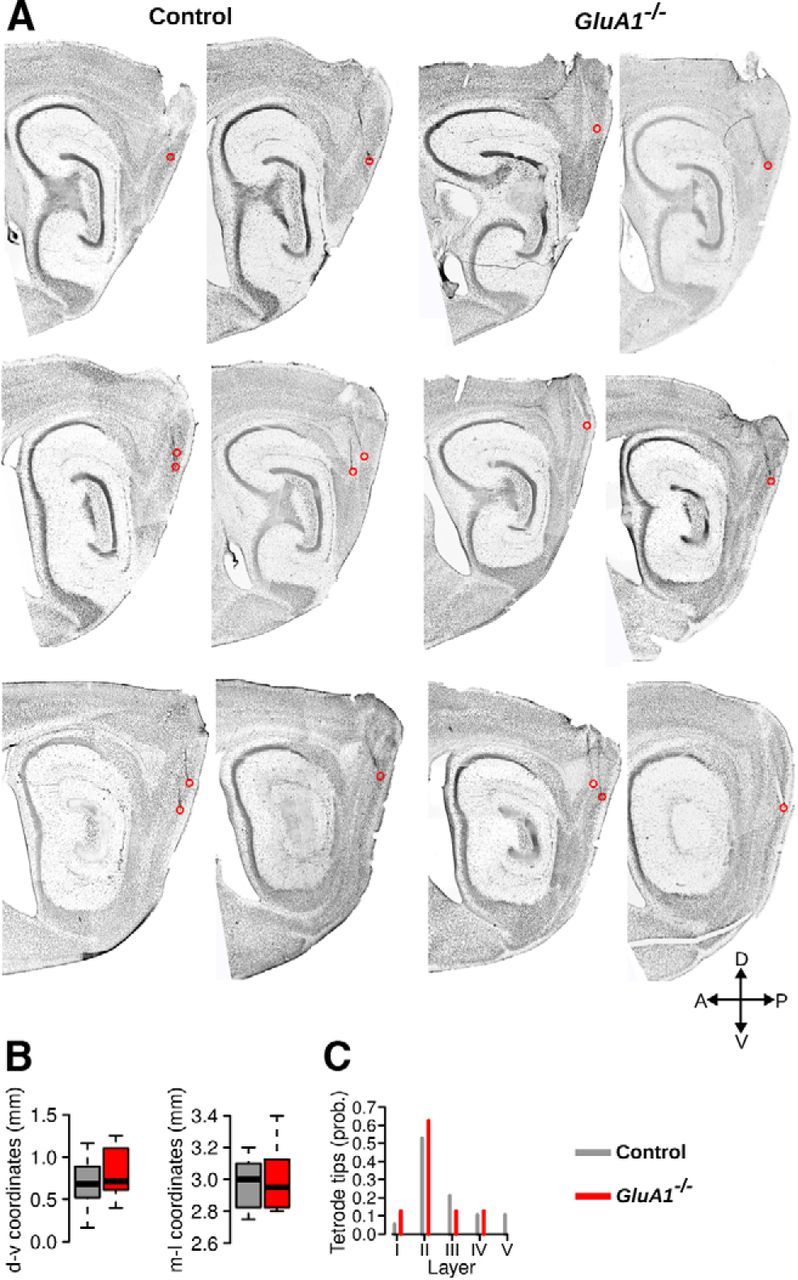
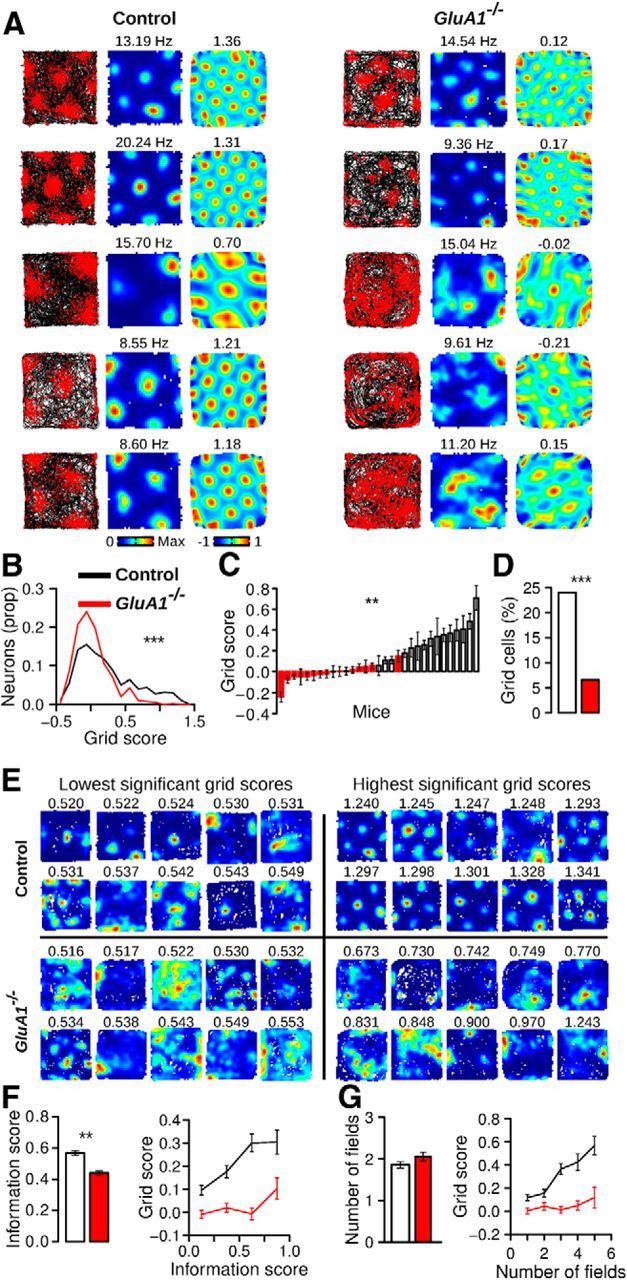
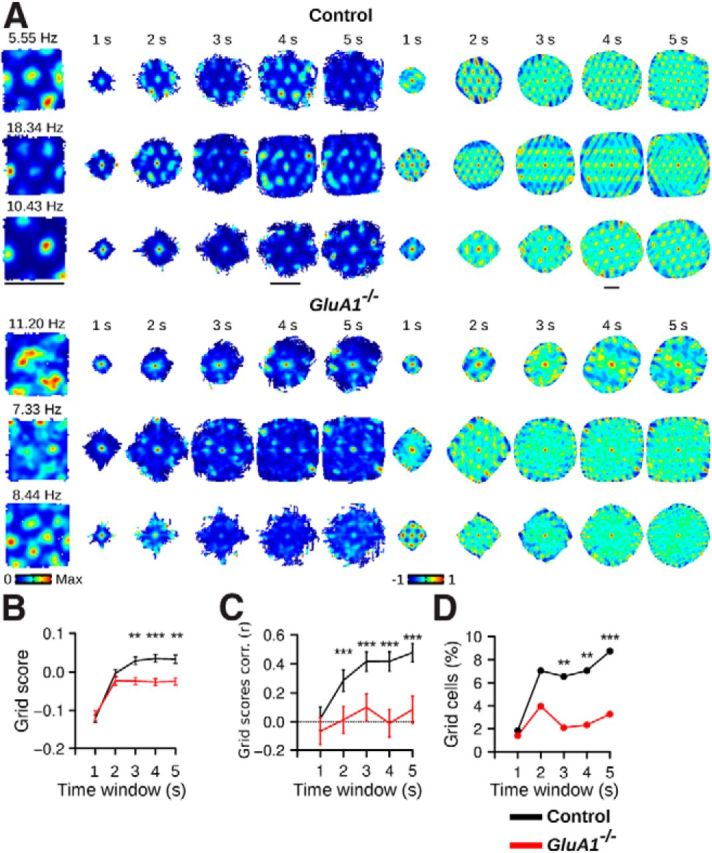
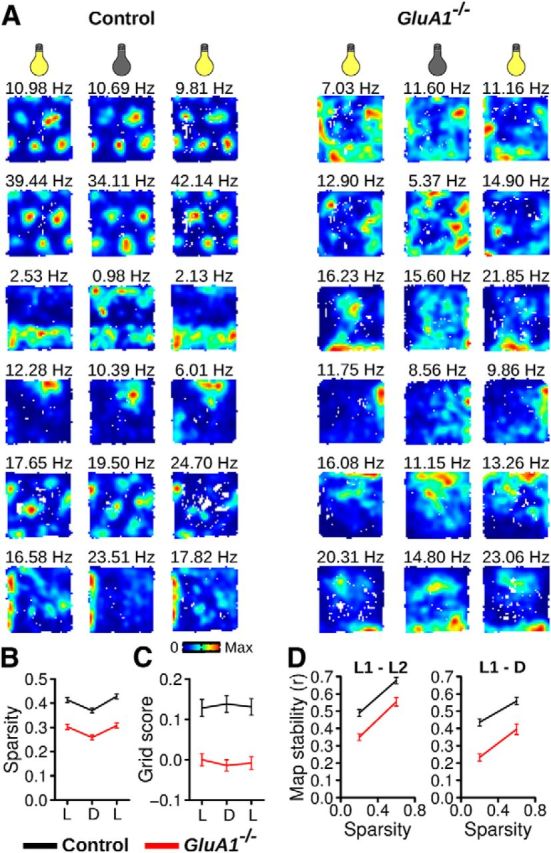
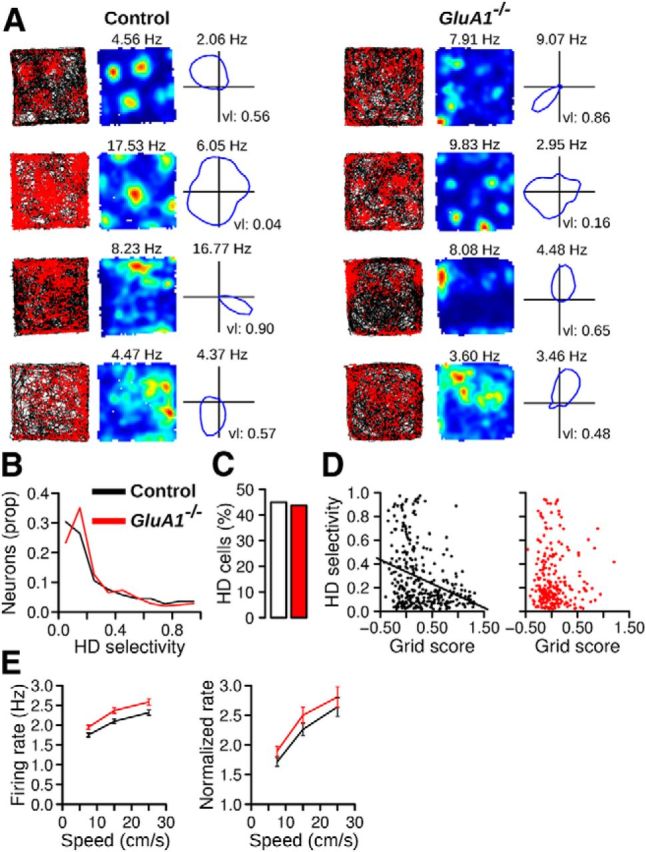
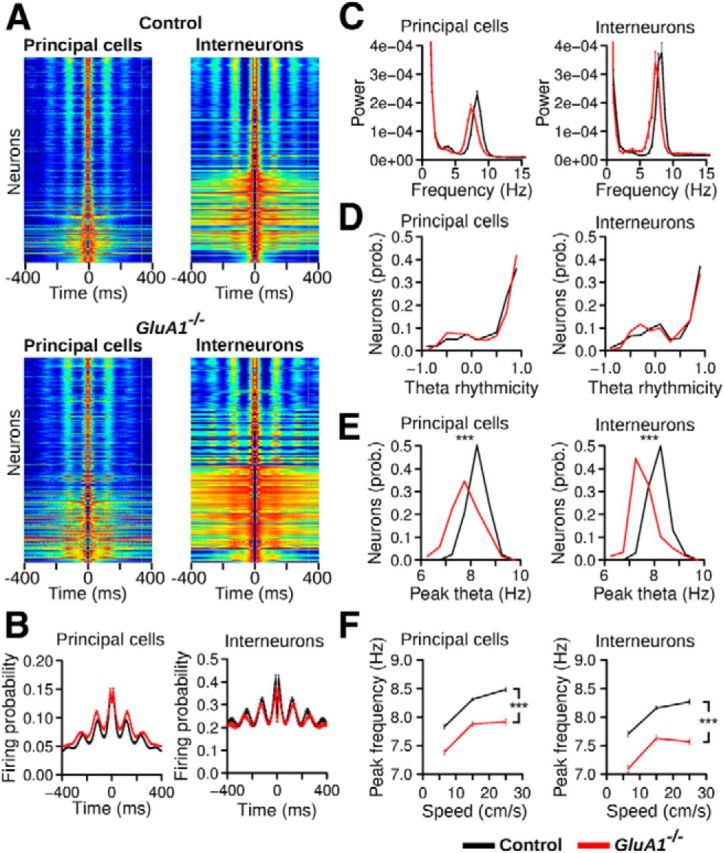
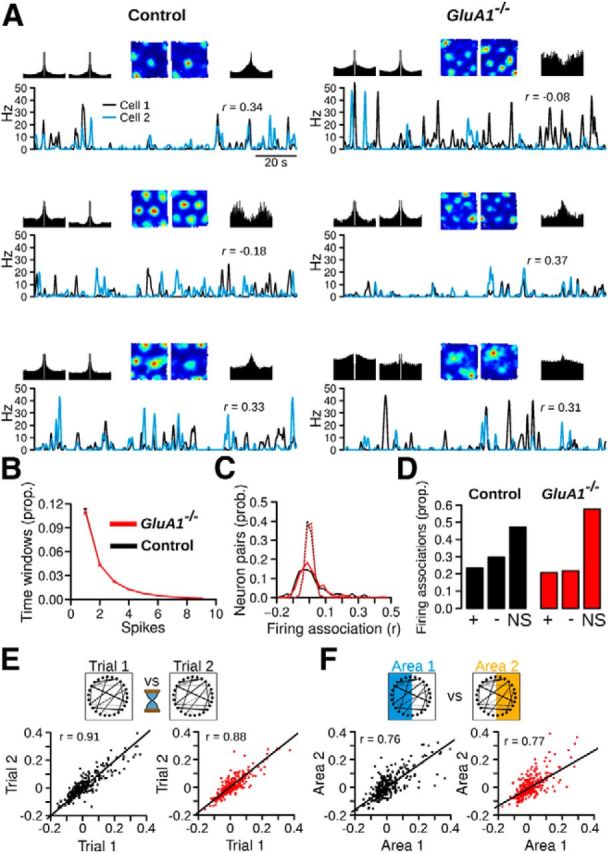
The hippocampus and the parahippocampal region have been proposed to contribute to path integration. Mice lacking GluA1-containing AMPA receptors (GluA1(-/-) mice) were previously shown to exhibit impaired hippocampal place cell selectivity. Here we investigated whether path integration performance and the activity of grid cells of the medial entorhinal cortex (MEC) are affected in these mice. We first tested GluA1(-/-) mice on a standard food-carrying homing task and found that they were impaired in processing idiothetic cues. To corroborate these findings, we developed an L-maze task that is less complex and is performed entirely in darkness, thereby reducing numerous confounding variables when testing path integration. Also in this task, the performance of GluA1(-/-) mice was impaired. Next, we performed in vivo recordings in the MEC of GluA1(-/-) mice. MEC neurons exhibited altered grid cell spatial periodicity and reduced spatial selectivity, whereas head direction tuning and speed modulation were not affected. The firing associations between pairs of neurons in GluA1(-/-) mice were stable, both in time and space, indicating that attractor states were still present despite the lack of grid periodicity. Together, these results support the hypothesis that spatial representations in the hippocampal-entorhinal network contribute to path integration.
Keywords: entorhinal cortex; glutamate receptors; grid cells; hippocampus; navigation; path integration.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases