

INO80 facilitates pluripotency gene activation in embryonic stem cell self-renewal, reprogramming, and blastocyst development
- PMID: 24792115
- PMCID: PMC4154226
- DOI: 10.1016/j.stem.2014.02.013
INO80 facilitates pluripotency gene activation in embryonic stem cell self-renewal, reprogramming, and blastocyst development
Abstract
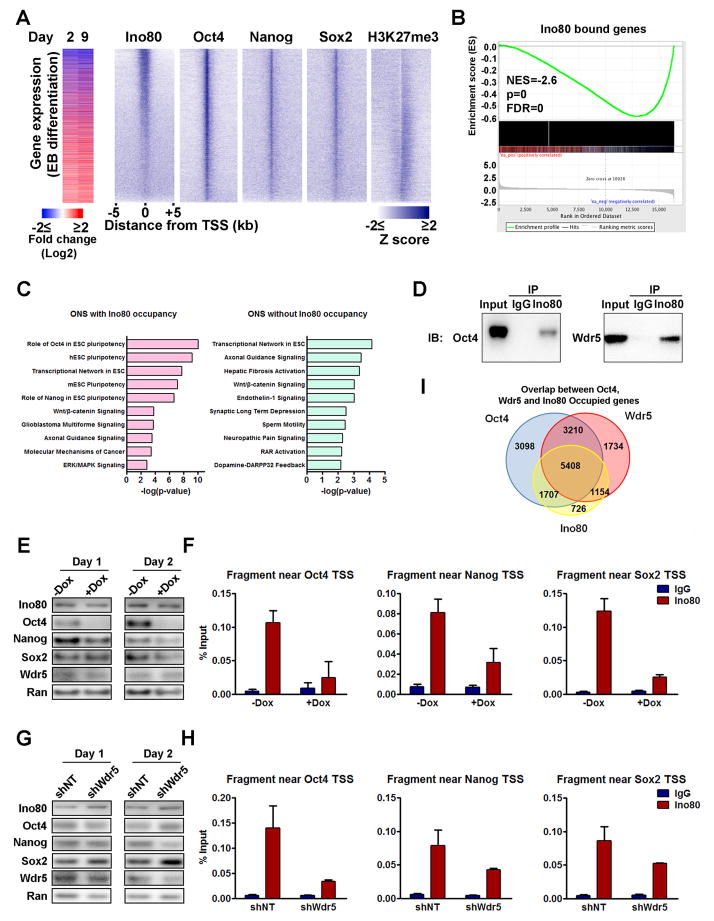
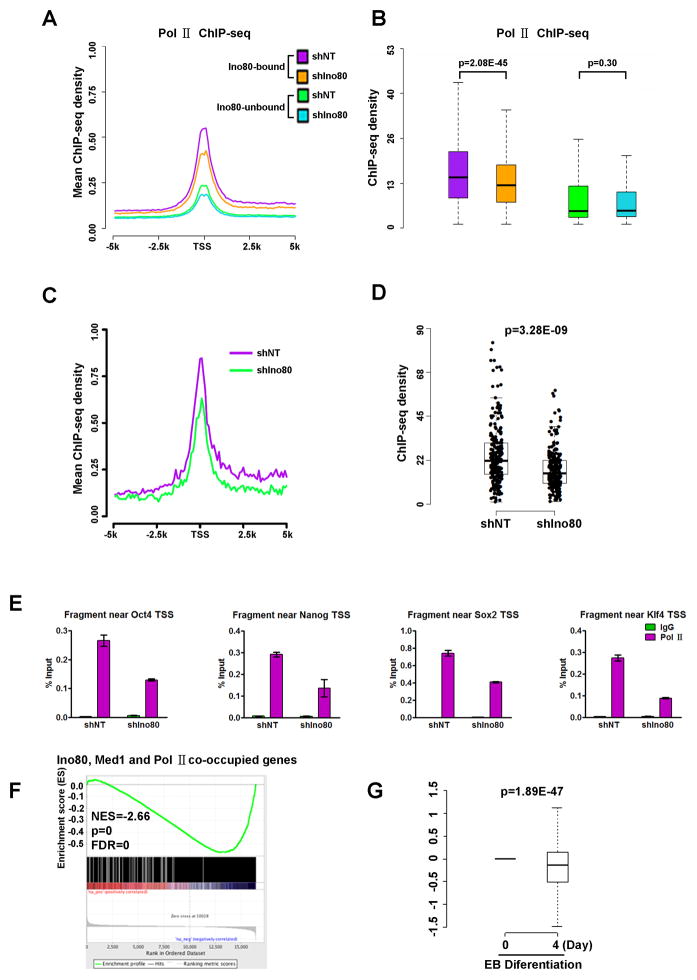
The master transcription factors play integral roles in the pluripotency transcription circuitry of embryonic stem cells (ESCs). How they selectively activate expression of the pluripotency network while simultaneously repressing genes involved in differentiation is not fully understood. Here, we define a requirement for the INO80 complex, a SWI/SNF family chromatin remodeler, in ESC self-renewal, somatic cell reprogramming, and blastocyst development. We show that Ino80, the chromatin remodeling ATPase, co-occupies pluripotency gene promoters with the master transcription factors, and its occupancy is dependent on OCT4 and WDR5. At the pluripotency genes, Ino80 maintains an open chromatin architecture and licenses recruitment of Mediator and RNA polymerase II for gene activation. Our data reveal an essential role for INO80 in the expression of the pluripotency network and illustrate the coordination among chromatin remodeler, transcription factor, and histone-modifying enzyme in the regulation of the pluripotent state.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures





Similar articles
-
SWI/SNF-Brg1 regulates self-renewal and occupies core pluripotency-related genes in embryonic stem cells.Stem Cells. 2009 Feb;27(2):317-28. doi: 10.1634/stemcells.2008-0710. Stem Cells. 2009. PMID: 19056910
-
[OCT4 and NANOG are the key genes in the system of pluripotency maintenance in mammalian cells].Genetika. 2008 Dec;44(12):1589-608. Genetika. 2008. PMID: 19178078 Review. Russian.
-
Reptin regulates pluripotency of embryonic stem cells and somatic cell reprogramming through Oct4-dependent mechanism.Stem Cells. 2014 Dec;32(12):3126-36. doi: 10.1002/stem.1827. Stem Cells. 2014. PMID: 25185564
-
Regulatory role of Klf5 in early mouse development and in embryonic stem cells.Vitam Horm. 2011;87:381-97. doi: 10.1016/B978-0-12-386015-6.00037-8. Vitam Horm. 2011. PMID: 22127252 Review.
-
Analysis of Oct4-dependent transcriptional networks regulating self-renewal and pluripotency in human embryonic stem cells.Stem Cells. 2007 Feb;25(2):500-10. doi: 10.1634/stemcells.2006-0426. Epub 2006 Oct 26. Stem Cells. 2007. PMID: 17068183
Cited by
-
Epigenome Regulation by Dynamic Nucleosome Unwrapping.Trends Biochem Sci. 2020 Jan;45(1):13-26. doi: 10.1016/j.tibs.2019.09.003. Epub 2019 Oct 17. Trends Biochem Sci. 2020. PMID: 31630896 Free PMC article. Review.
-
LncRNA HAND2-AS1 promotes liver cancer stem cell self-renewal via BMP signaling.EMBO J. 2019 Sep 2;38(17):e101110. doi: 10.15252/embj.2018101110. Epub 2019 Jul 23. EMBO J. 2019. PMID: 31334575 Free PMC article.
-
Long Noncoding RNAs and Cancer Stem Cells: Dangerous Liaisons Managing Cancer.Int J Mol Sci. 2023 Jan 17;24(3):1828. doi: 10.3390/ijms24031828. Int J Mol Sci. 2023. PMID: 36768150 Free PMC article. Review.
-
ATP-Dependent Chromatin Remodellers in Inner Ear Development.Cells. 2023 Feb 7;12(4):532. doi: 10.3390/cells12040532. Cells. 2023. PMID: 36831199 Free PMC article. Review.
-
Ino80 is required for H2A.Z eviction from hypha-specific promoters and hyphal development of Candida albicans.Mol Microbiol. 2022 Jul;118(1-2):92-104. doi: 10.1111/mmi.14954. Epub 2022 Jun 28. Mol Microbiol. 2022. PMID: 35713098 Free PMC article.
References
-
- Chen X, Xu H, Yuan P, Fang F, Huss M, Vega VB, Wong E, Orlov YL, Zhang W, Jiang J, et al. Integration of external signaling pathways with the core transcriptional network in embryonic stem cells. Cell. 2008;133:1106–1117. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
