

Mouse spermatogenic stem cells continually interconvert between equipotent singly isolated and syncytial states
- PMID: 24792118
- PMCID: PMC4010676
- DOI: 10.1016/j.stem.2014.01.019
Mouse spermatogenic stem cells continually interconvert between equipotent singly isolated and syncytial states
Abstract
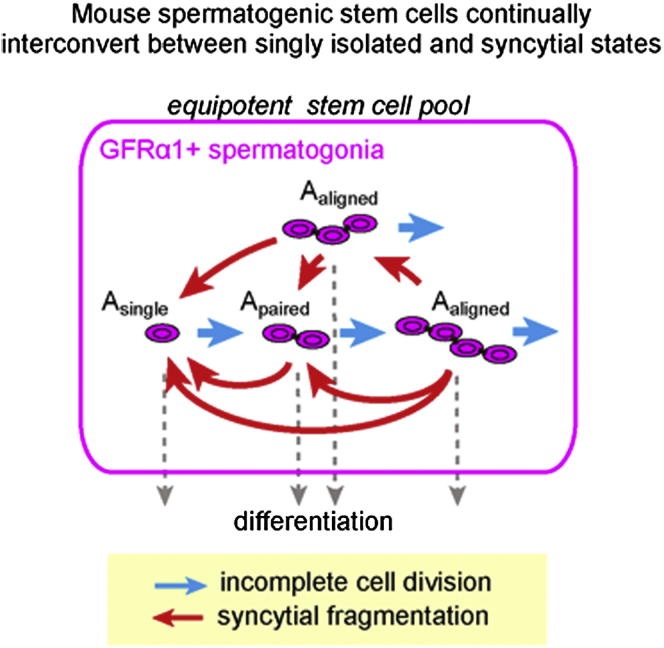
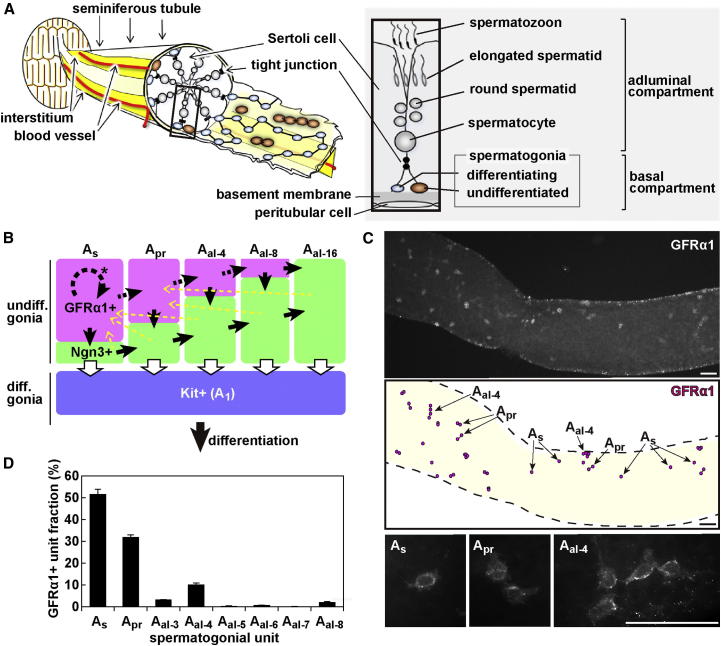
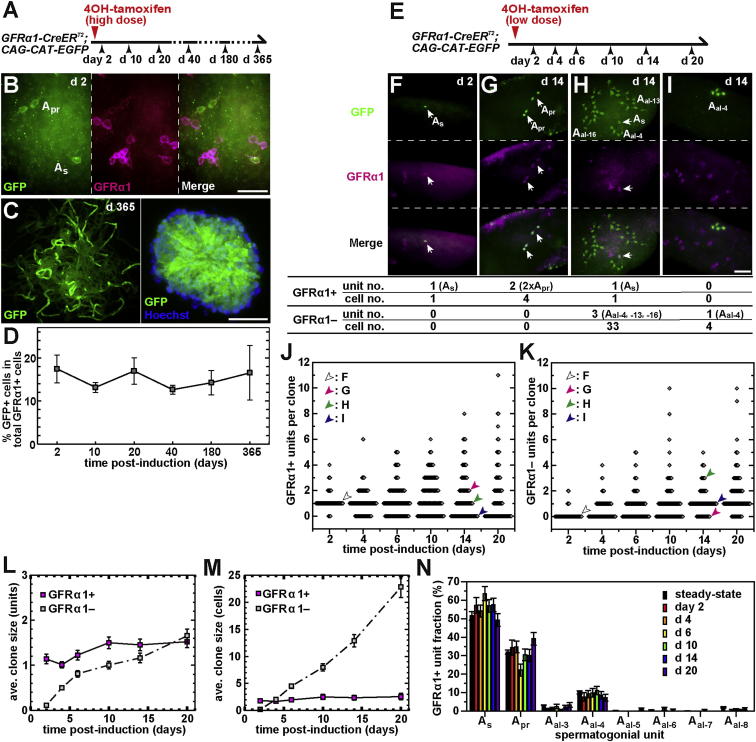
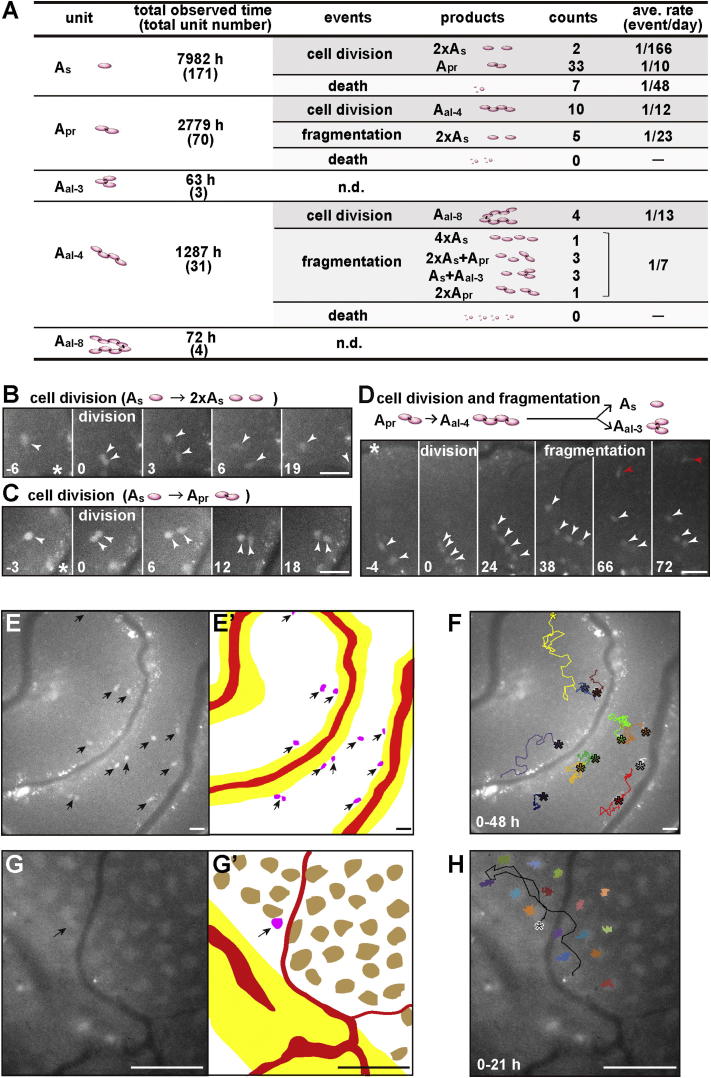
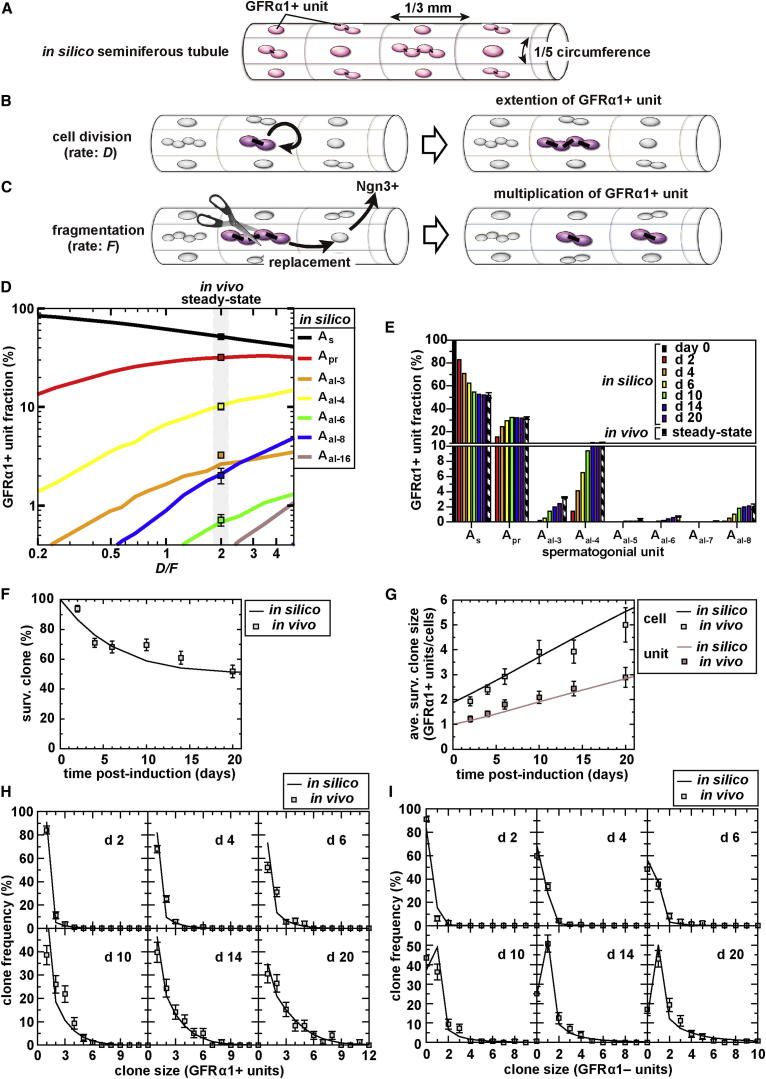
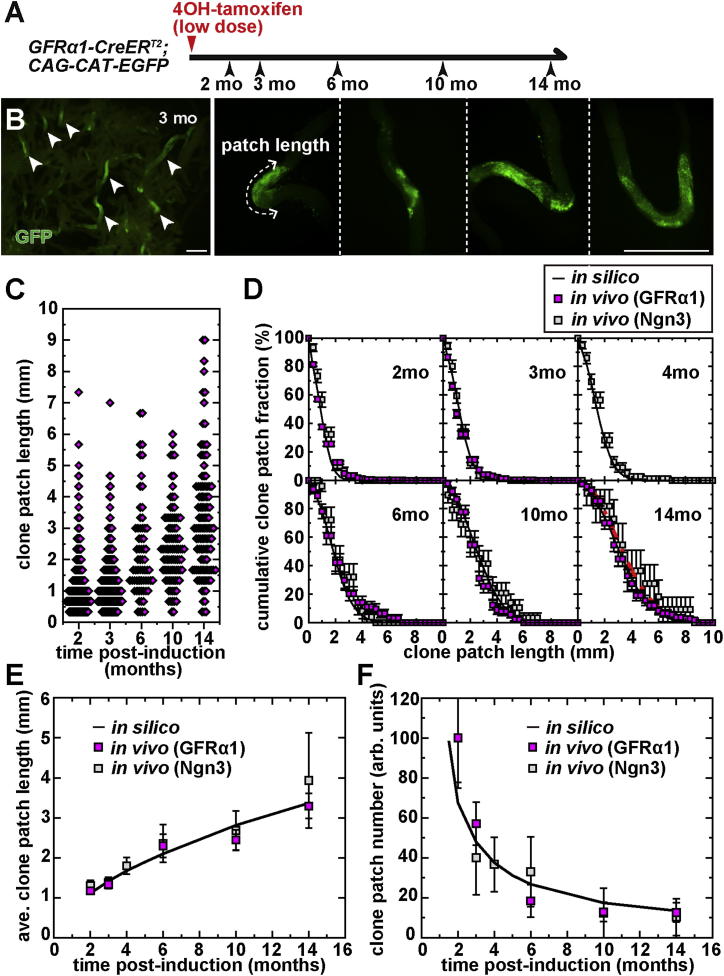
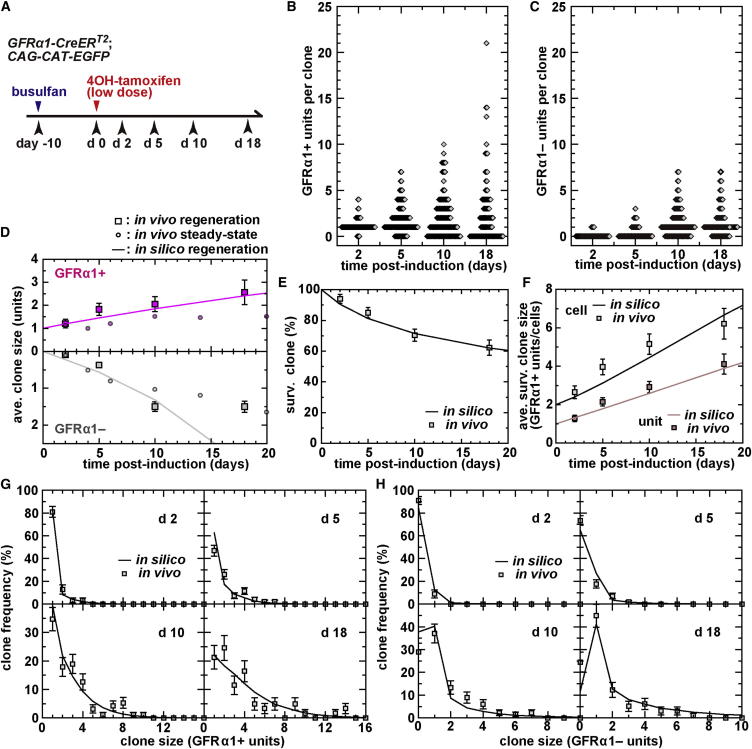
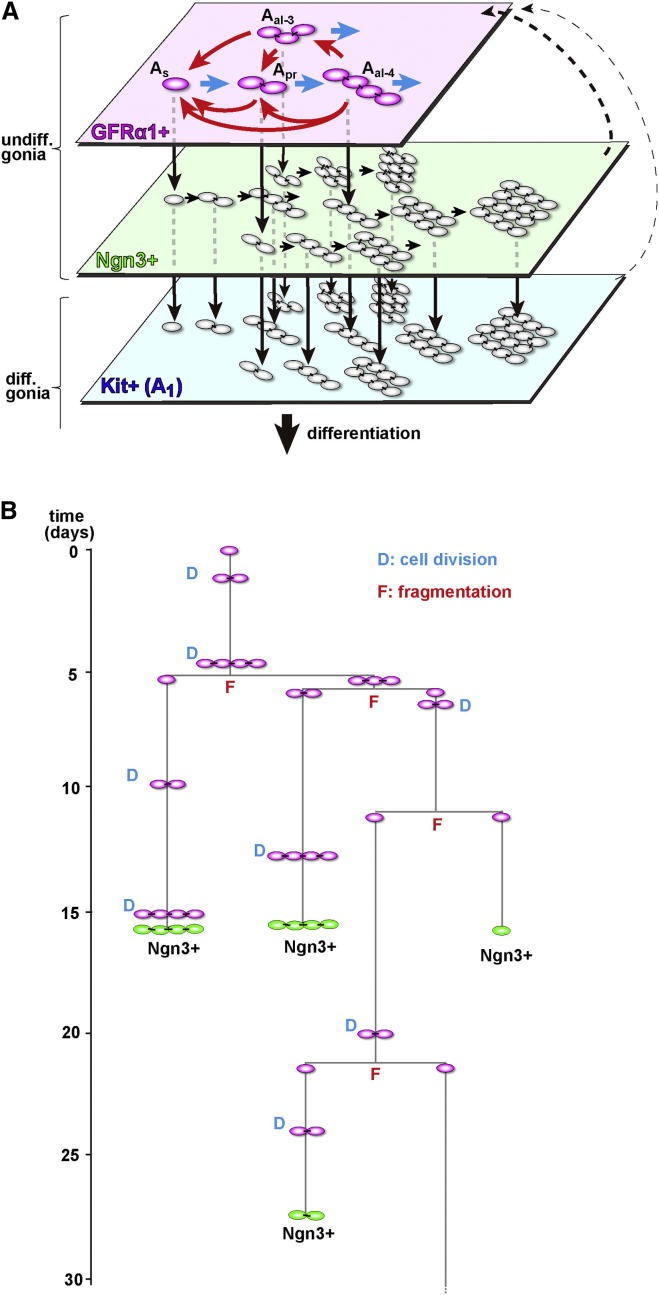
The identity and behavior of mouse spermatogenic stem cells have been a long-standing focus of interest. In the prevailing "As model," stem cell function is restricted to singly isolated (As) spermatogonia. By examining single-cell dynamics of GFRα1+ stem cells in vivo, we evaluate an alternative hypothesis that, through fragmentation, syncytial spermatogonia also contribute to stem cell function in homeostasis. We use live imaging and pulse labeling to quantitatively determine the fates of individual GFRα1+ cells and find that, during steady-state spermatogenesis, the entire GFRα1+ population comprises a single stem cell pool, in which cells continually interconvert between As and syncytial states. A minimal biophysical model, relying only on the rates of incomplete cell division and syncytial fragmentation, precisely predicts the stochastic fates of GFRα1+ cells during steady state and postinsult regeneration. Thus, our results define an alternative and dynamic model for spermatogenic stem cell function in the mouse testis.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Barroca V., Lassalle B., Coureuil M., Louis J.P., Le Page F., Testart J., Allemand I., Riou L., Fouchet P. Mouse differentiating spermatogonia can generate germinal stem cells in vivo. Nat. Cell Biol. 2009;11:190–196. - PubMed
-
- Braun R.E., Behringer R.R., Peschon J.J., Brinster R.L., Palmiter R.D. Genetically haploid spermatids are phenotypically diploid. Nature. 1989;337:373–376. - PubMed
-
- Chiarini-Garcia H., Hornick J.R., Griswold M.D., Russell L.D. Distribution of type A spermatogonia in the mouse is not random. Biol. Reprod. 2001;65:1179–1185. - PubMed
-
- de Rooij D.G., Russell L.D. All you wanted to know about spermatogonia but were afraid to ask. J. Androl. 2000;21:776–798. - PubMed
-
- Erickson B.H. Survival and renewal of murine stem spermatogonia following 60Co gamma radiation. Radiat. Res. 1981;86:34–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
