

Rigidity sensing and adaptation through regulation of integrin types
- PMID: 24793358
- PMCID: PMC4031069
- DOI: 10.1038/nmat3960
Rigidity sensing and adaptation through regulation of integrin types
Abstract
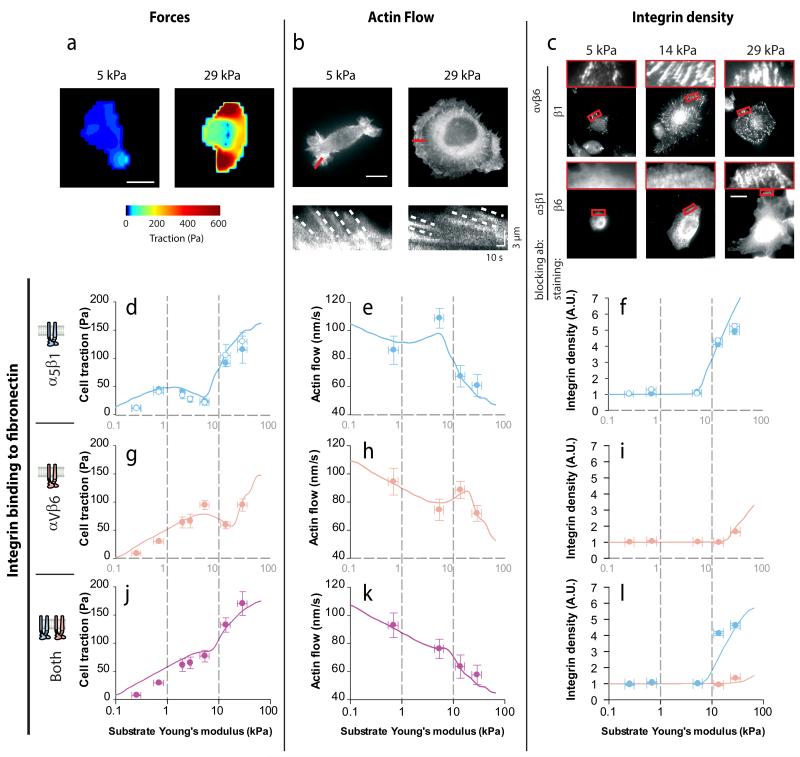
Tissue rigidity regulates processes in development, cancer and wound healing. However, how cells detect rigidity, and thereby modulate their behaviour, remains unknown. Here, we show that sensing and adaptation to matrix rigidity in breast myoepithelial cells is determined by the bond dynamics of different integrin types. Cell binding to fibronectin through either α5β1 integrins (constitutively expressed) or αvβ6 integrins (selectively expressed in cancer and development) adapts force generation, actin flow and integrin recruitment to rigidities associated with healthy or malignant tissue, respectively. In vitro experiments and theoretical modelling further demonstrate that this behaviour is explained by the different binding and unbinding rates of both integrin types to fibronectin. Moreover, rigidity sensing through differences in integrin bond dynamics applies both when integrins bind separately and when they compete for binding to fibronectin.
Figures



Comment in
-
Cellular mechanotransduction: sensing rigidity.Nat Mater. 2014 Jun;13(6):539-40. doi: 10.1038/nmat3996. Nat Mater. 2014. PMID: 24845988 No abstract available.
References
-
- Wang HB, Dembo M, Wang YL. Substrate flexibility regulates growth and apoptosis of normal but not transformed cells. Am J Physiol Cell Physiol. 2000;279(5):C1345–1350. - PubMed
-
- Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell. 2006;126(4):677–689. - PubMed
-
- Paszek MJ, Zahir N, Johnson KR, Lakins JN, Rozenberg GI, Gefen A, et al. Tensional homeostasis and the malignant phenotype. Cancer Cell. 2005;8(3):241–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
