

Ecdysone mediates the development of immunity in the Drosophila embryo
- PMID: 24794300
- PMCID: PMC4030305
- DOI: 10.1016/j.cub.2014.03.062
Ecdysone mediates the development of immunity in the Drosophila embryo
Abstract
Beyond their role in cell metabolism, development, and reproduction, hormones are also important modulators of the immune system. In the context of inflammatory disorders, systemic administration of pharmacological doses of synthetic glucocorticoids (GCs) is widely used as an anti-inflammatory treatment [1, 2]. However, not all actions of GCs are immunosuppressive, and many studies have suggested that physiological concentrations of GCs can have immunoenhancing effects [3-7]. For a more comprehensive understanding of how steroid hormones regulate immunity and inflammation, a simple in vivo system is required. The Drosophila embryo has recently emerged as a powerful model system to study the recruitment of immune cells to sterile wounds [8] and host-pathogen dynamics [9]. Here we investigate the immune response of the fly embryo to bacterial infections and find that the steroid hormone 20-hydroxyecdysone (20-HE) can regulate the quality of the immune response and influence the resolution of infection in Drosophila embryos.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
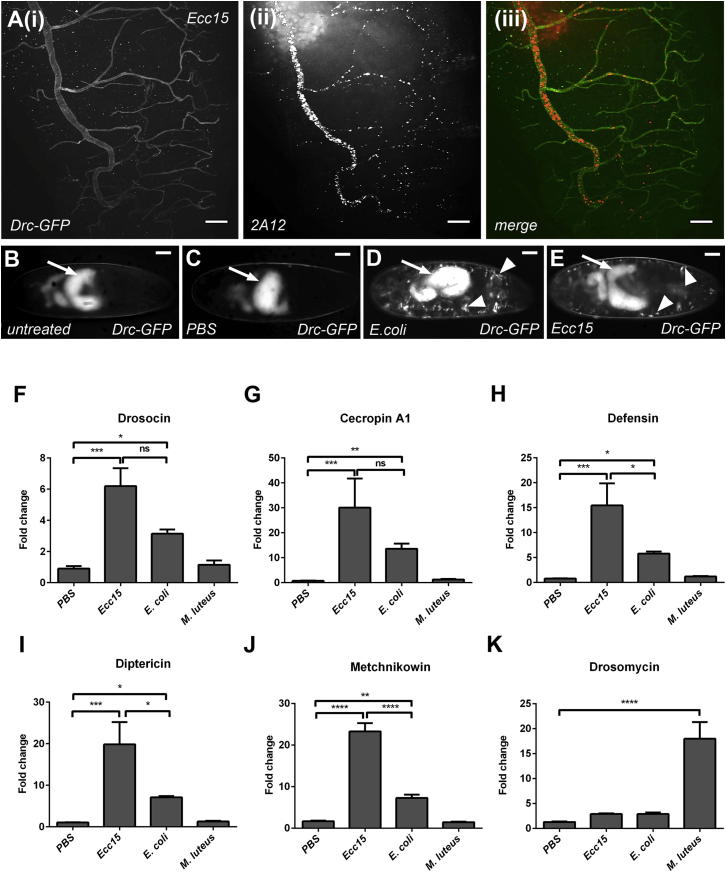
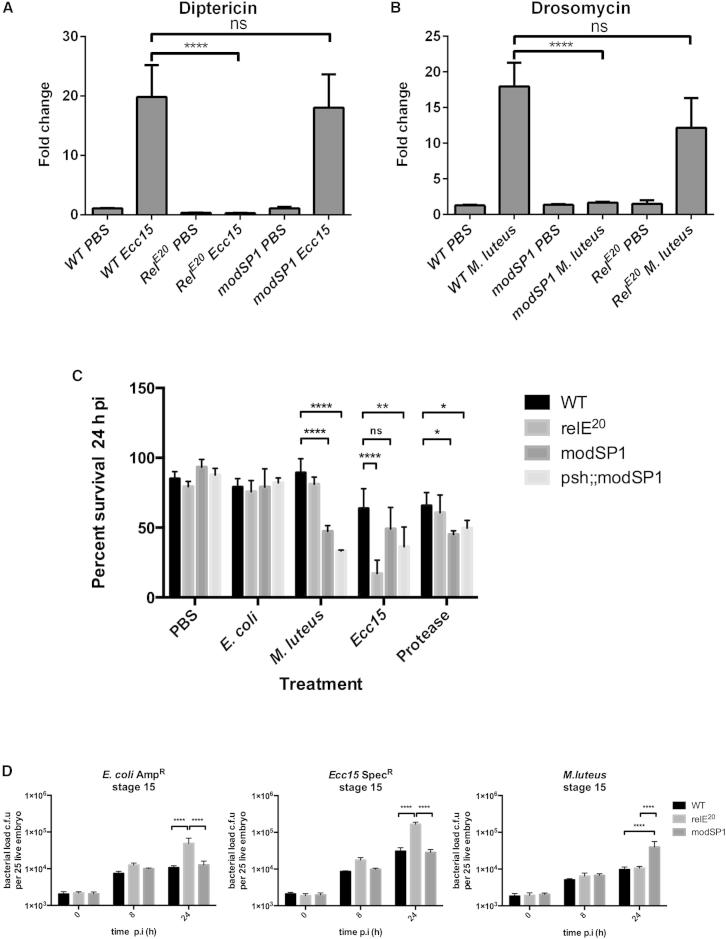
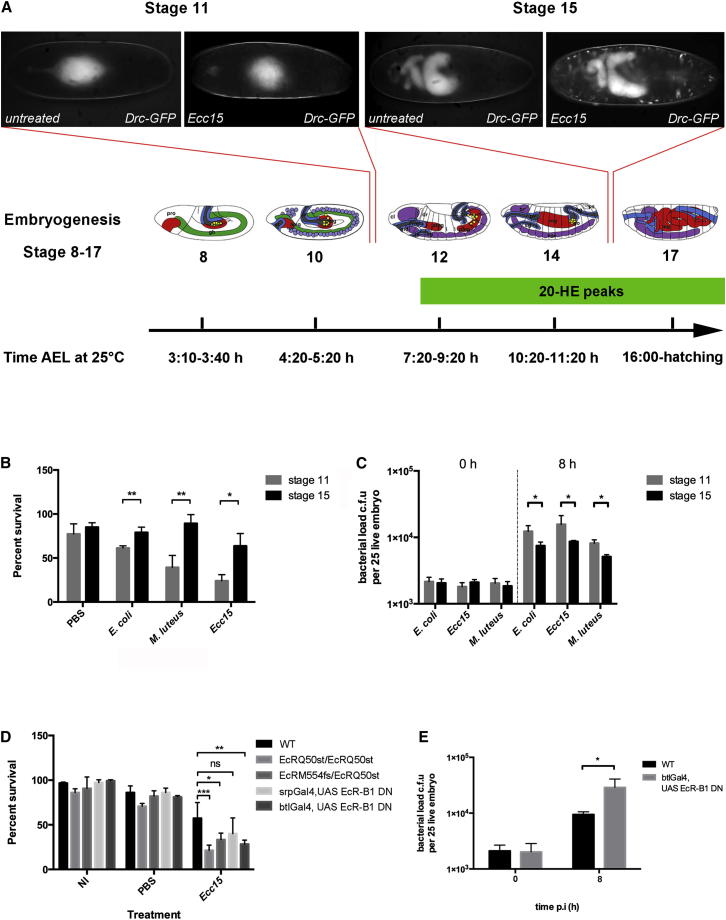
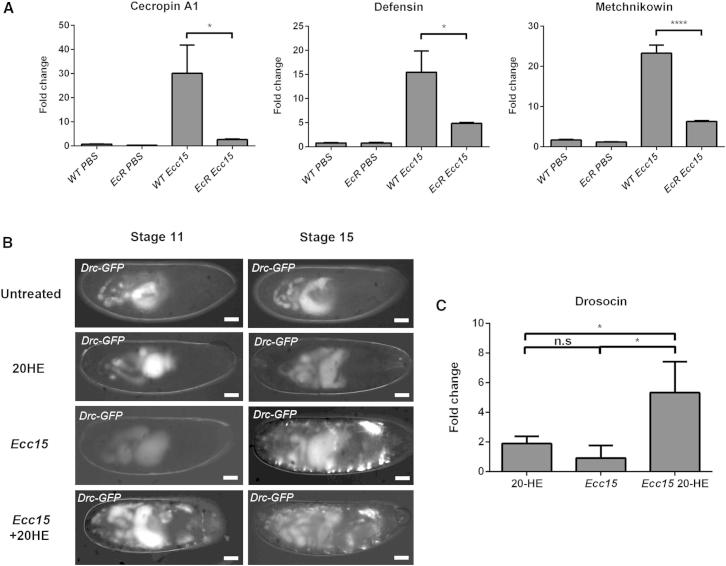
References
-
- Necela B.M., Cidlowski J.A. Mechanisms of glucocorticoid receptor action in noninflammatory and inflammatory cells. Proc. Am. Thorac. Soc. 2004;1:239–246. - PubMed
-
- Galon J., Franchimont D., Hiroi N., Frey G., Boettner A., Ehrhart-Bornstein M., O’Shea J.J., Chrousos G.P., Bornstein S.R. Gene profiling reveals unknown enhancing and suppressive actions of glucocorticoids on immune cells. FASEB J. 2002;16:61–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
