

Review
doi: 10.1016/j.dnarep.2014.03.029.
Epub 2014 Apr 30.
Ribonucleotides in DNA: origins, repair and consequences
Affiliations
- PMID: 24794402
- PMCID: PMC4065383
- DOI: 10.1016/j.dnarep.2014.03.029
Item in Clipboard
Review
Ribonucleotides in DNA: origins, repair and consequences
DNA Repair (Amst).
2014 Jul.
Abstract
While primordial life is thought to have been RNA-based (Cech, Cold Spring Harbor Perspect. Biol. 4 (2012) a006742), all living organisms store genetic information in DNA, which is chemically more stable. Distinctions between the RNA and DNA worlds and our views of "DNA" synthesis continue to evolve as new details emerge on the incorporation, repair and biological effects of ribonucleotides in DNA genomes of organisms from bacteria through humans.
Keywords: DNA polymerase; DNA repair; DNA replication; Genome instability; Ribonucleotides.
Published by Elsevier B.V.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures
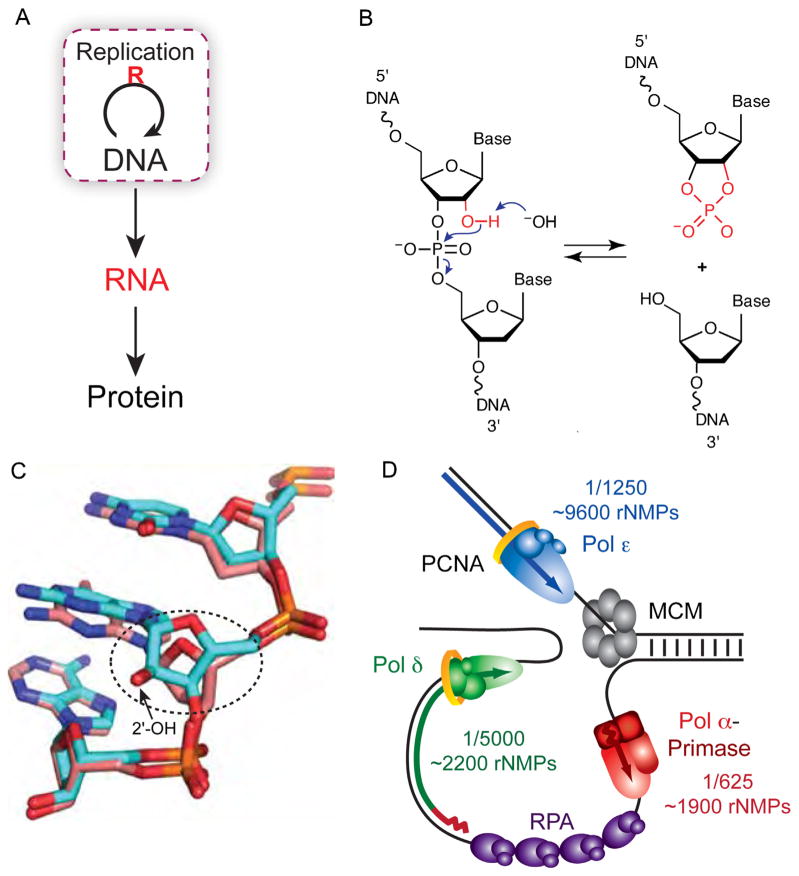
Ribonucleotides in DNA. (A) The central dogma of molecular biology describes how genetic information is transferred [1]. The essential loop has been modified to highlight the discovery that ribonucleotides are incorporated into DNA during replication. (B) Hydrolysis of the DNA backbone occurs at the site of a ribonucleotide in the presence of alkali when the deprotonated 2′ hydroxyl of a ribonucleotide cleaves the DNA strand via nucleophilic attack of the phosphate backbone. The resulting 2′–3′ cyclic phosphate is opened in the presence of water to yield equimolar amounts of 2′ and 3′ phosphate (not shown). (C) The Dickerson dodecamer NMR solution structure from [7] containing all DNA (dd-DNA, coral) or a single riboG (rG4-DNA, cyan). Shown is the view looking into the minor groove. Encircled is the alteration in sugar pucker from C2′-endo in the dd-DNA structure to C3′-endo in the rG4-DNA structure (Fig. 3 in [7]). (D) Model of the yeast replication fork with the frequency and estimated number of ribonucleotides incorporated by each DNA polymerase (adapted from Fig. 2D in [31]).
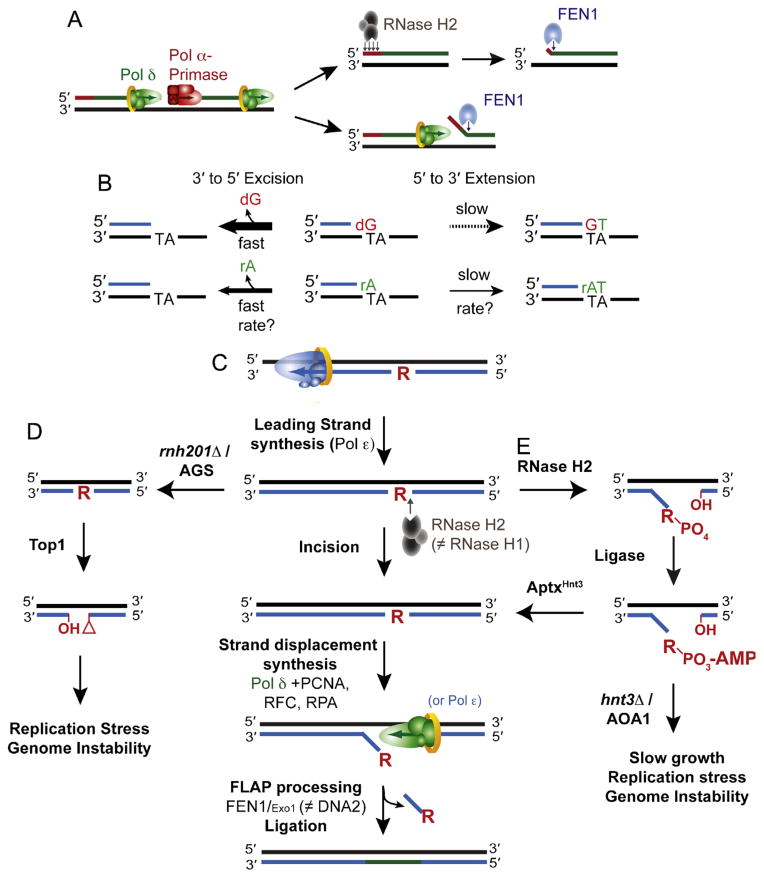
Origins and removal of ribonucleotides in DNA. (A) Incorporation and removal of ribonucleotides incorporated by RNA primase during OF synthesis and maturation. Two of the pathways by which the RNA primer synthesized by Pol α-primase is removed are displayed. RNase H2 is able to digest and remove the RNA primer up until the terminal 3′ ribonucleotide that is removed by FEN1. Alternatively, Pol δ can perform strand displacement synthesis to generate a flap that is degraded by FEN1. (B) Proofreading efficiency is determined by the balance between excision and extension. The polymerase exonuclease activity can excise ribonucleotides from the 3′ terminus in primer-template DNA, an event that is more efficient on a mismatched 3′ terminal dNMP primer than it is on an rNMP. The kinetics of proofreading (both excision and extension) by a replicative DNA polymerase in the presence of a terminal rNMP remain to be biochemically determined. (C) The ribonucleotide excision repair (RER) pathway is initiated by incision 5′ to the ribonucleotide by RNase H2. Strand displacement synthesis by the polymerase is followed by nucleolytic flap processing and ligation. There are redundant functions of the polymerases (Pol δ and Polε) as well as the nucleases (FEN1 and Exo1; Adapted from Fig. 4 in [56]). (D) Loss of RNase H2 activity due to deletion of RNH201 or RNase H2-deficiency in AGS leaves unrepaired genomic ribonucleotides that may become targets for Top1-cleavage and removal. The resulting unligatable DNA ends possess a 2′–3′-cyclic phosphate and a 5′-hydroxyl that may promote the formation of DNA breaks and/or recombination and cause genome instability. (Adapted from Fig. 4 in [60]). (E) The pathway by which RNase H2 cleavage at ribonucleotides can generate abortive ligation intermediates (adenylated 5′-RNA–DNA junctions) that require APTXHNT3 for resolution. Failure of this pathway due to deletion of HNT3 or APTX deficiency in AOA1 causes phenotypes that may reflect the persistence of adenylated RNA–DNA junctions (Adapted from Fig. 2E in [57].
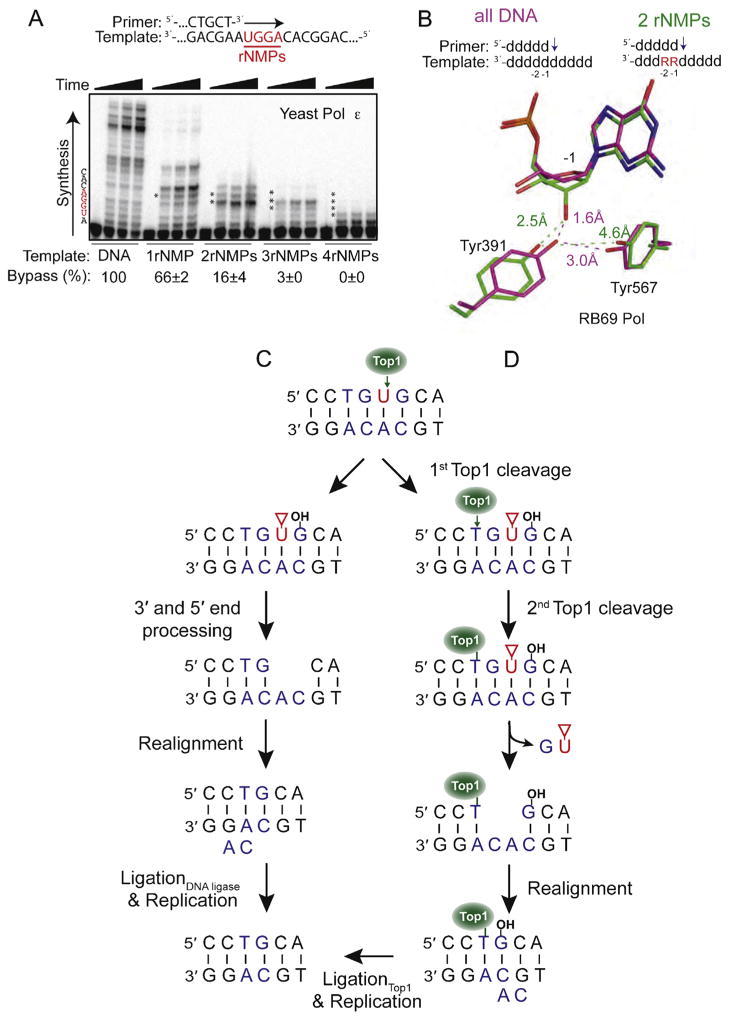
Consequences of ribonucleotides in DNA. (A) Bypass efficiency of ribonucleotides by yeast Pol ε. Primer extension was performed using five different primer-templates that varied in the number of template ribonucleotides (shown in red). Bypass efficiency (%) was calculated relative to the all-DNA control (Adapted from Fig. 1B in [73]). (B) The presence of two consecutive ribonucleotides in a template affects sugar pucker and positioning of two conserved active site tyrosine residues in RB69 Pol. The positioning of Tyr391 and Tyr567 in the presence of all-DNA template (magenta) is compared to the positions of these residues in the presence of a ribonucleotide at the −1 position (green). (Adapted from Fig. 4 in [73]). (C) The first of two models for generation of a two base deletion following Top1 cleavage at a ribonucleotide. In this pathway, end processing of the damaged 3′ and 5′ ends generates a gap within the repeat unit. Following misalignment of the complementary strands, ligation and replication cause loss of one repeat unit (Adapted from Fig. 4 in [59]). The red triangle denotes a 2′–3′-cyclic phosphate. (D) The second model involves sequential Top1-cleavages for generation of an rNMP-associated deletion. Following the first incision, there is a second Top1-cleavage reaction on the 5′ side of the nick that causes release of the DNA between the two cleavage sites and trapping of Top1. Realignment of the complementary DNA strands moves the free 5′-OH in close proximity to Top1 so that it can catalyze religation (Adapted from Fig. 6 in [83]).
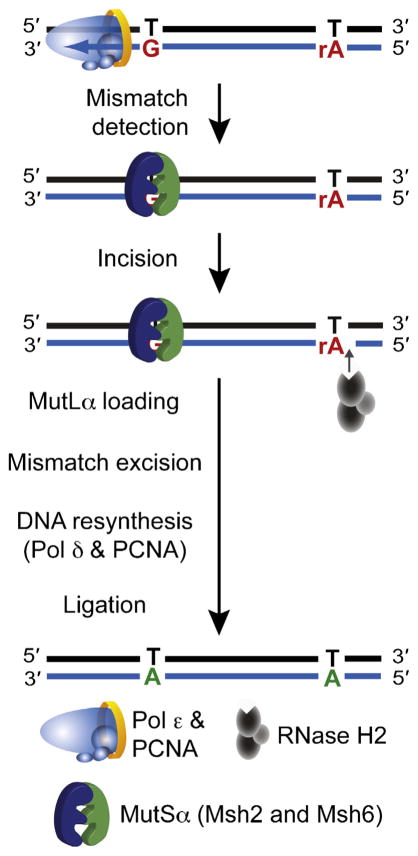
A model depicting how ribonucleotides incorporated by Pol ε act as a signal for MMR of errors generated in nascent leading strand DNA. Following recognition of the mismatch by the MutSα heterodimer, incision 5′ to a ribonucleotide by RNase H2 creates a nick that can be used as a loading site for MutLα. This nick can be distant from the site of the mismatch, because MutLα is able to translocate along the DNA and incise at a site close to the mismatch to allow excision, resynthesis and ligation.
References
-
- Crick F. Central dogma of molecular biology. Nature. 1970;227:561–563. - PubMed
-
- Li Y, Breaker RR. Kinetics of RNA degradation by specific base catalysis of transesterification involving the 2′-hydroxyl group. J Am Chem Soc. 1999;121:5326–5372.
-
- Egli M, Usman N, Rich A. Conformational influence of the ribose 2′-hydroxyl group: crystal structures of DNA–RNA chimeric duplexes. Biochemistry. 1993;32:3221–3237. - PubMed
-
- Jaishree TN, van der Marel GA, van Boom JH, Wang AH. Structural influence of RNA incorporation in DNA: quantitative nuclear magnetic resonance refinement of d(CG)r(CG)d(CG) and d(CG)r(C)d(TAGCG) Biochemistry. 1993;32:4903–4911. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
