

The terminal oxidase cbb3 functions in redox control of magnetite biomineralization in Magnetospirillum gryphiswaldense
- PMID: 24794567
- PMCID: PMC4097590
- DOI: 10.1128/JB.01652-14
The terminal oxidase cbb3 functions in redox control of magnetite biomineralization in Magnetospirillum gryphiswaldense
Abstract
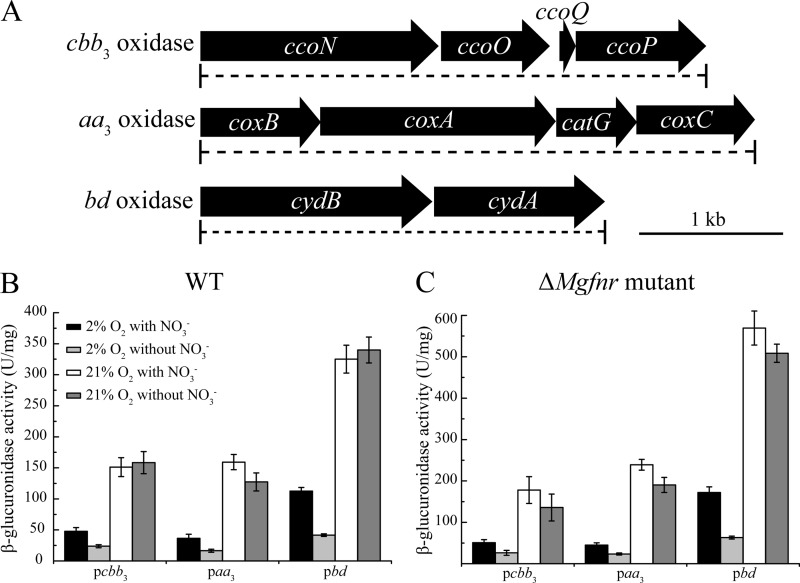
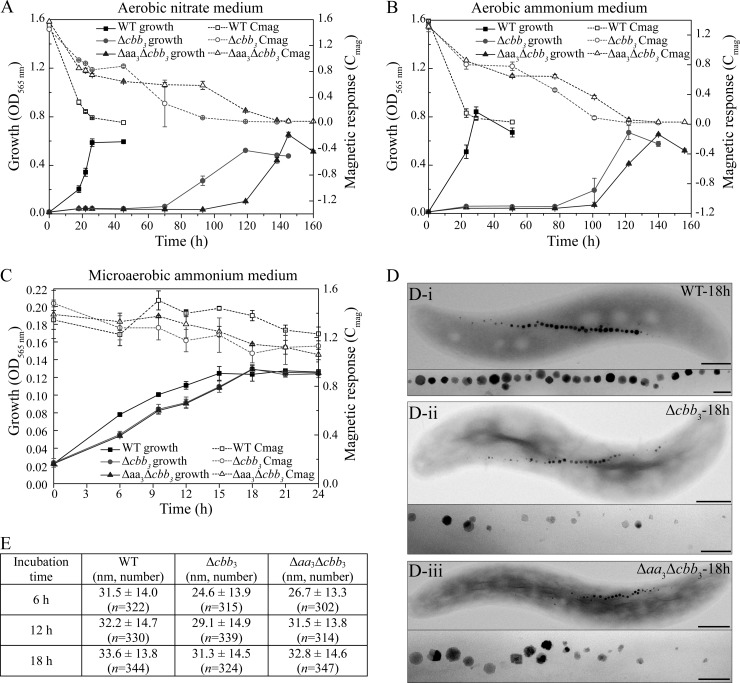
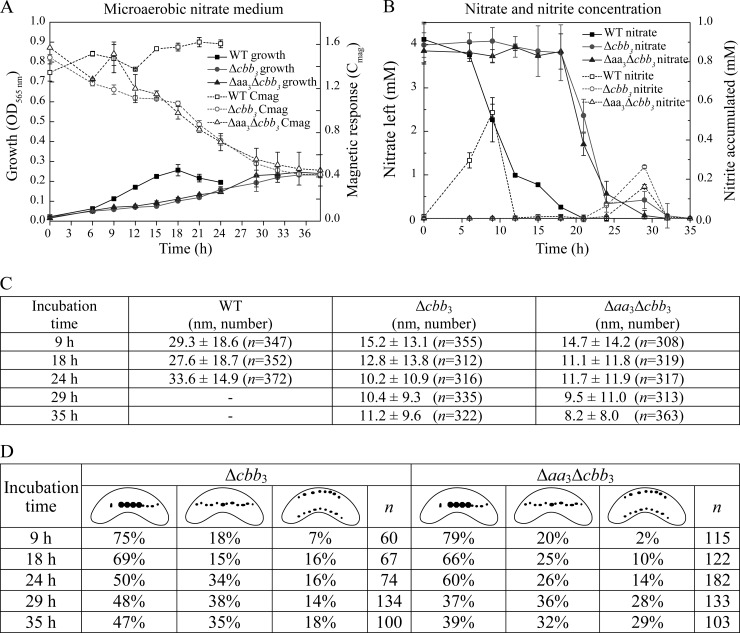
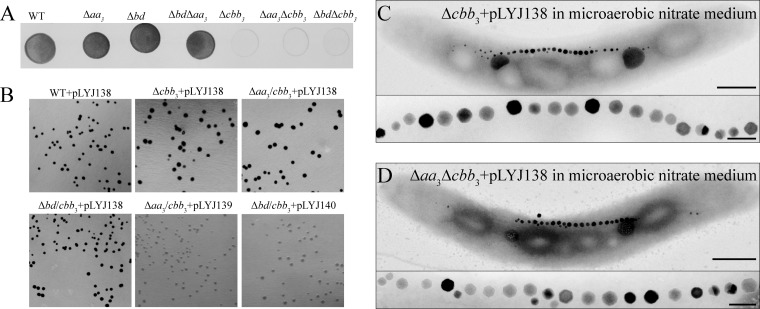
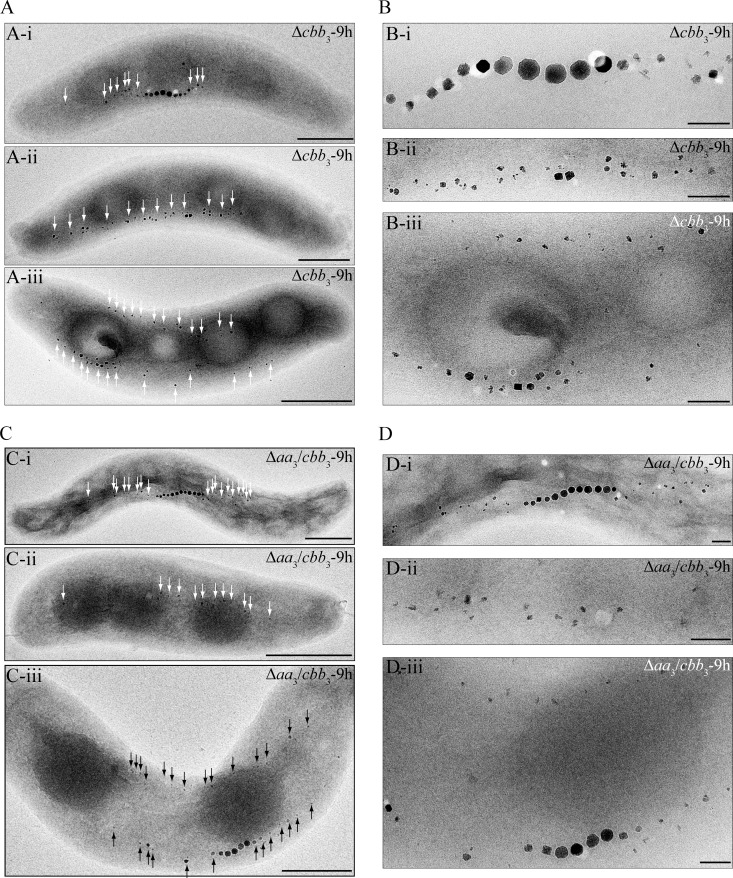
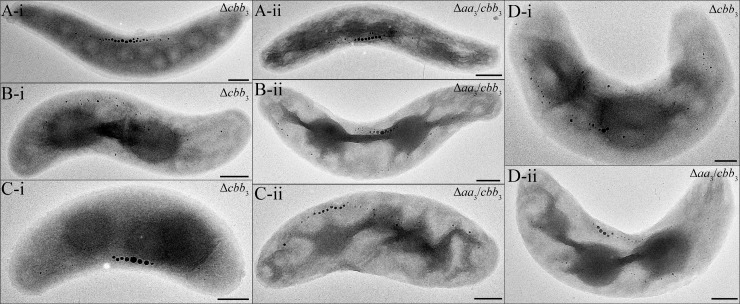
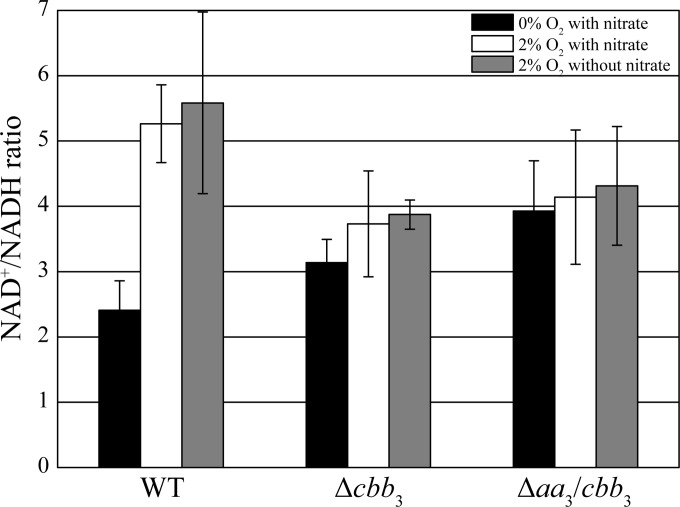
The biomineralization of magnetosomes in Magnetospirillum gryphiswaldense and other magnetotactic bacteria occurs only under suboxic conditions. However, the mechanism of oxygen regulation and redox control of biosynthesis of the mixed-valence iron oxide magnetite [FeII(FeIII)2O4] is still unclear. Here, we set out to investigate the role of aerobic respiration in both energy metabolism and magnetite biomineralization of M. gryphiswaldense. Although three operons encoding putative terminal cbb3-type, aa3-type, and bd-type oxidases were identified in the genome assembly of M. gryphiswaldense, genetic and biochemical analyses revealed that only cbb3 and bd are required for oxygen respiration, whereas aa3 had no physiological significance under the tested conditions. While the loss of bd had no effects on growth and magnetosome synthesis, inactivation of cbb3 caused pleiotropic effects under microaerobic conditions in the presence of nitrate. In addition to their incapability of simultaneous nitrate and oxygen reduction, cbb3-deficient cells had complex magnetosome phenotypes and aberrant morphologies, probably by disturbing the redox balance required for proper growth and magnetite biomineralization. Altogether, besides being the primary terminal oxidase for aerobic respiration, cbb3 oxidase may serve as an oxygen sensor and have a further role in poising proper redox conditions required for magnetite biomineralization.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Schübbe S, Kube M, Scheffel A, Wawer C, Heyen U, Meyerdierks A, Madkour MH, Mayer F, Reinhardt R, Schüler D. 2003. Characterization of a spontaneous nonmagnetic mutant of Magnetospirillum gryphiswaldense reveals a large deletion comprising a putative magnetosome island. J. Bacteriol. 185:5779–5790. 10.1128/JB.185.19.5779-5790.2003 - DOI - PMC - PubMed
-
- Ullrich S, Kube M, Schübbe S, Reinhardt R, Schüler D. 2005. A hypervariable 130-kilobase genomic region of Magnetospirillum gryphiswaldense comprises a magnetosome island which undergoes frequent rearrangements during stationary growth. J. Bacteriol. 187:7176–7184. 10.1128/JB.187.21.7176-7184.2005 - DOI - PMC - PubMed
-
- Lohsse A, Ullrich S, Katzmann E, Borg S, Wanner G, Richter M, Voigt B, Schweder T, Schüler D. 2011. Functional analysis of the magnetosome island in Magnetospirillum gryphiswaldense: the mamAB operon is sufficient for magnetite biomineralization. PLoS One 6:e25561. 10.1371/journal.pone.0025561 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
