

Dysfunction of spatacsin leads to axonal pathology in SPG11-linked hereditary spastic paraplegia
- PMID: 24794856
- PMCID: PMC4140466
- DOI: 10.1093/hmg/ddu200
Dysfunction of spatacsin leads to axonal pathology in SPG11-linked hereditary spastic paraplegia
Abstract
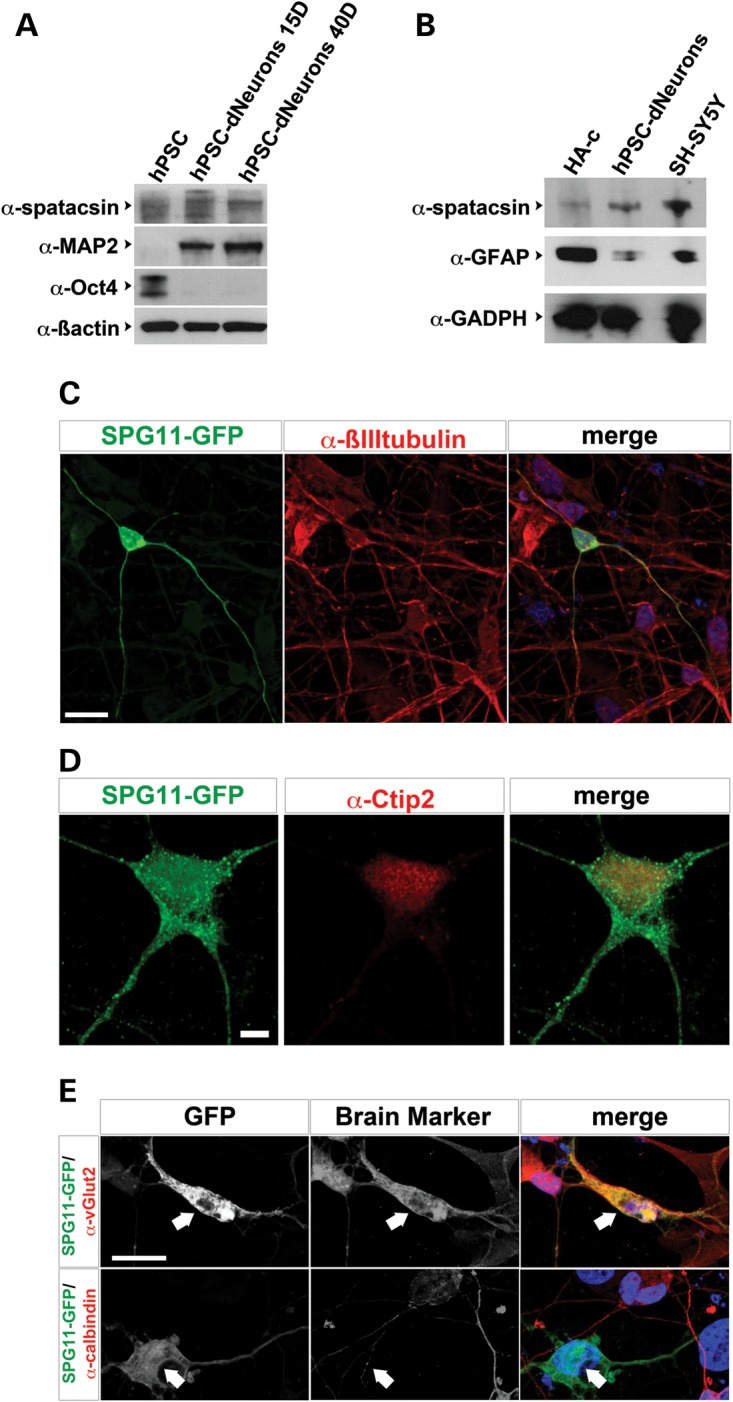
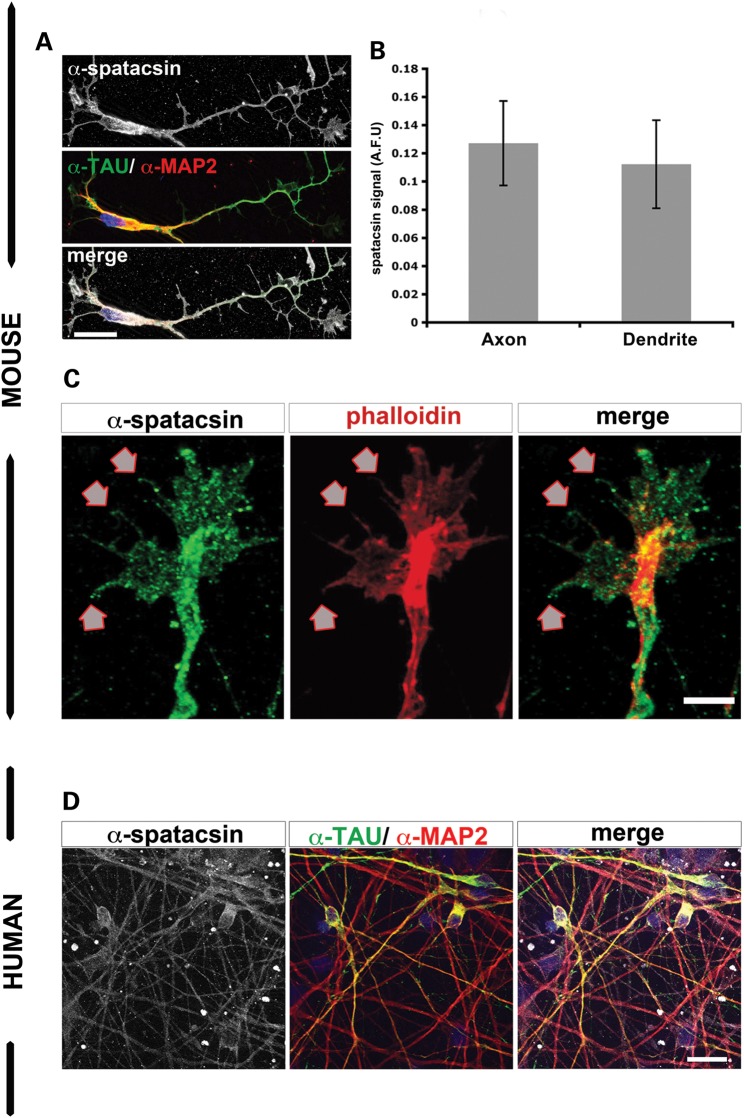
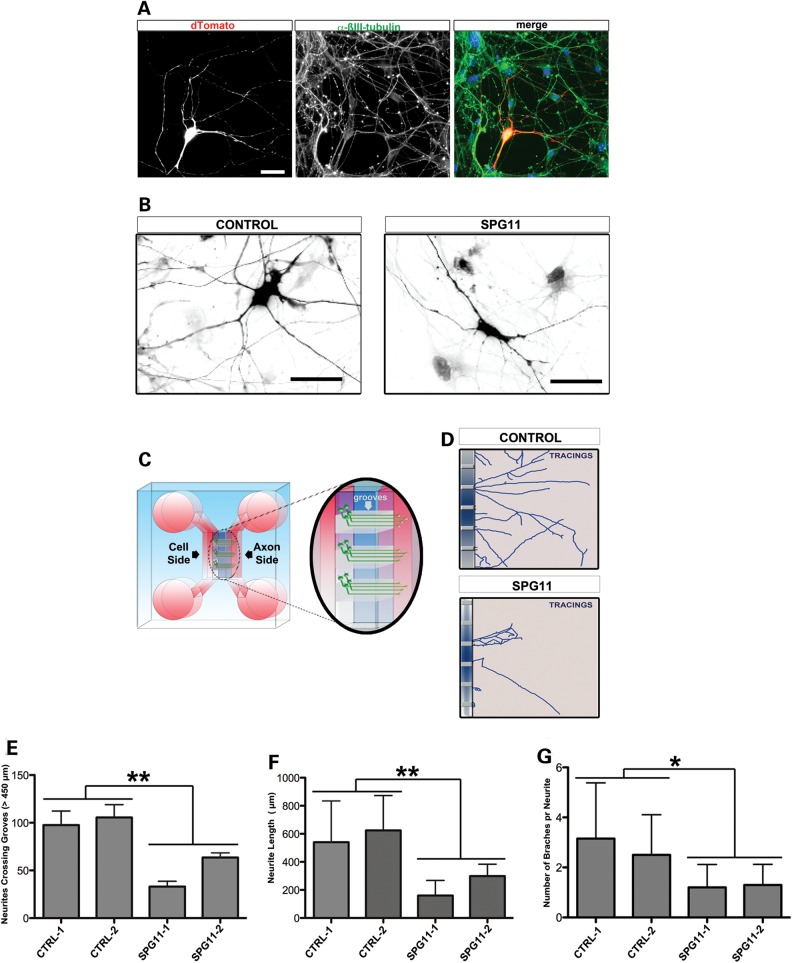
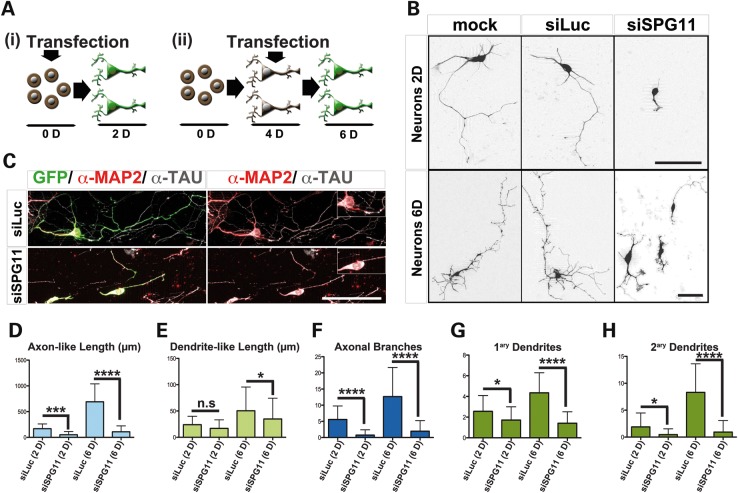
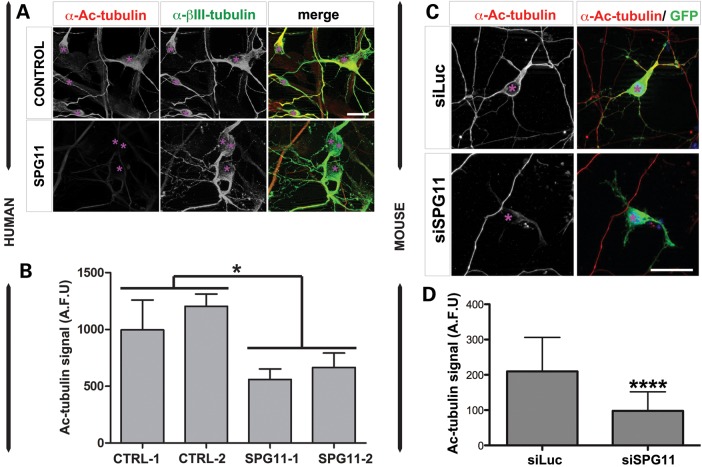
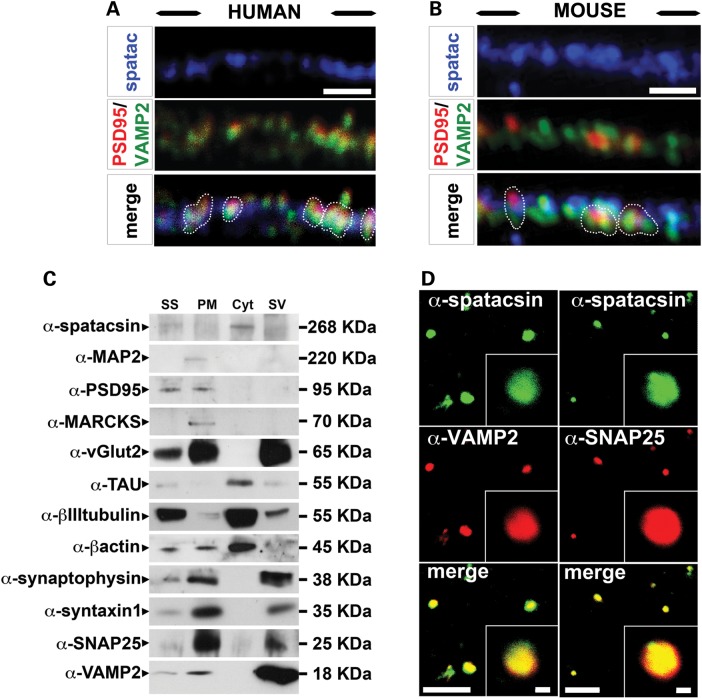
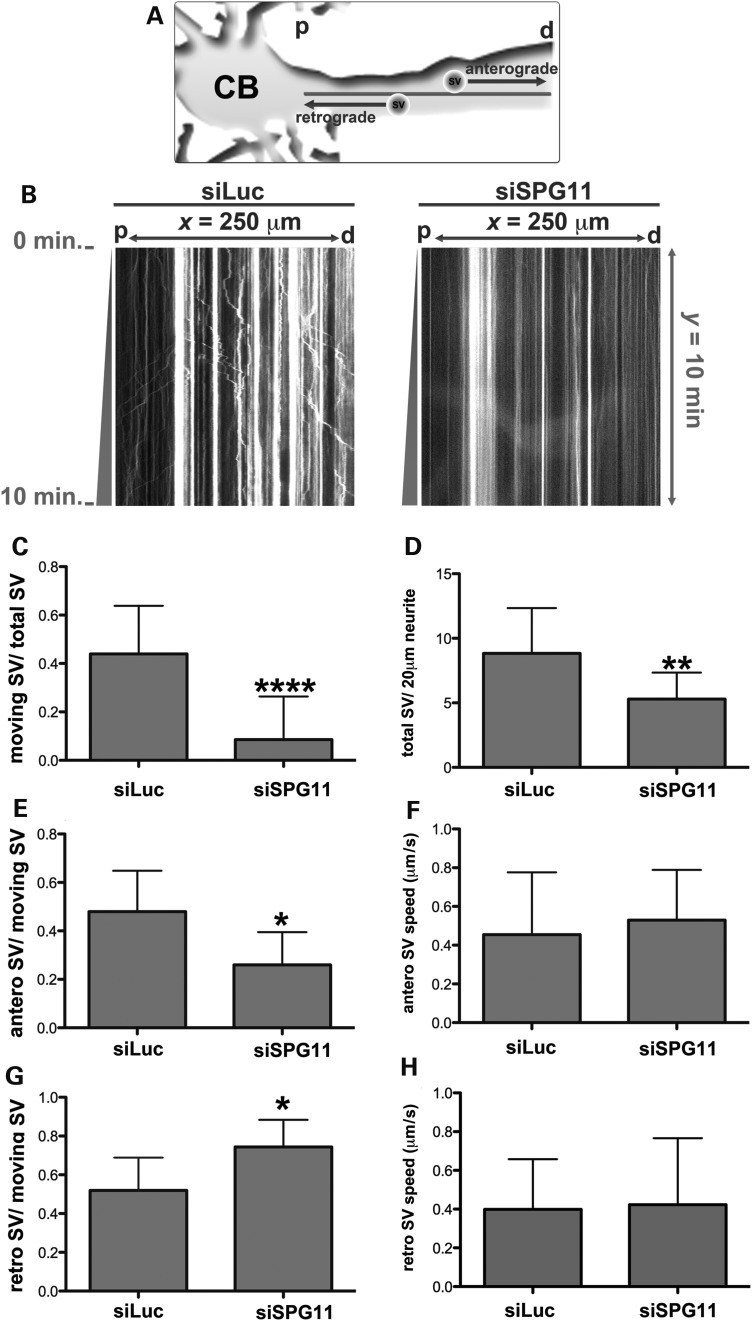
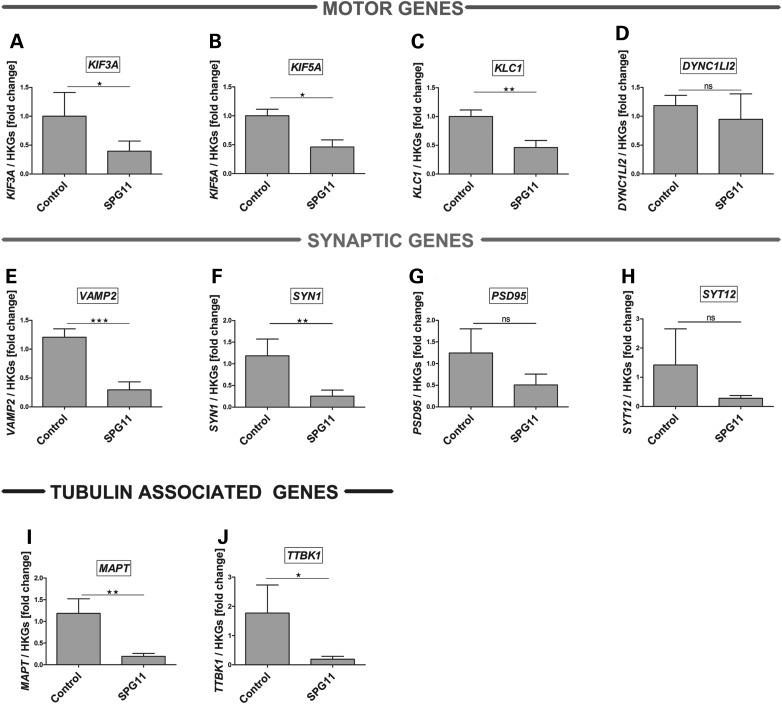
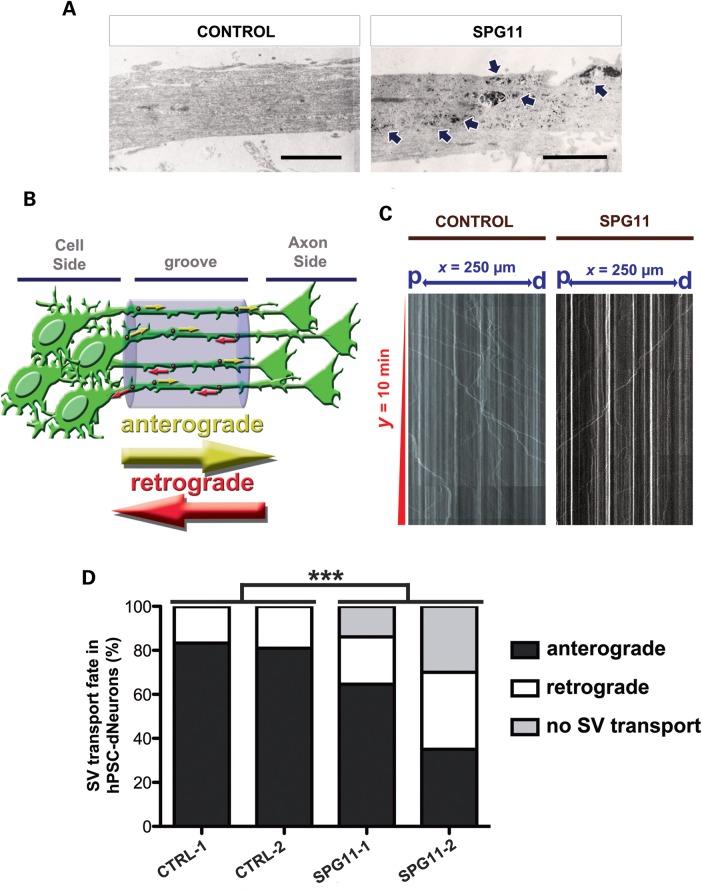
Hereditary spastic paraplegias are a group of inherited motor neuron diseases characterized by progressive paraparesis and spasticity. Mutations in the spastic paraplegia gene SPG11, encoding spatacsin, cause an autosomal-recessive disease trait; however, the precise knowledge about the role of spatacsin in neurons is very limited. We for the first time analyzed the expression and function of spatacsin in human forebrain neurons derived from human pluripotent stem cells including lines from two SPG11 patients and two controls. SPG11 patients'-derived neurons exhibited downregulation of specific axonal-related genes, decreased neurite complexity and accumulation of membranous bodies within axonal processes. Altogether, these data point towards axonal pathologies in human neurons with SPG11 mutations. To further corroborate spatacsin function, we investigated human pluripotent stem cell-derived neurons and mouse cortical neurons. In these cells, spatacsin was located in axons and dendrites. It colocalized with cytoskeletal and synaptic vesicle (SV) markers and was present in synaptosomes. Knockdown of spatacsin in mouse cortical neurons evidenced that the loss of function of spatacsin leads to axonal instability by downregulation of acetylated tubulin. Finally, time-lapse assays performed in SPG11 patients'-derived neurons and spatacsin-silenced mouse neurons highlighted a reduction in the anterograde vesicle trafficking indicative of impaired axonal transport. By employing SPG11 patient-derived forebrain neurons and mouse cortical neurons, this study provides the first evidence that SPG11 is implicated in axonal maintenance and cargo trafficking. Understanding the cellular functions of spatacsin will allow deciphering mechanisms of motor cortex dysfunction in autosomal-recessive hereditary spastic paraplegia.
© The Author 2014. Published by Oxford University Press.
Figures
References
-
- Harding A.E. Classification of the hereditary ataxias and paraplegias. Lancet. 1983;1:1151–1155. - PubMed
-
- Bettencourt C., Lopez-Sendon J., Garcia-Caldentey J., Rizzu P., Bakker I., Shomroni O., Quintans B., Davila J., Bevova M., Sobrido M.J., et al. Exome sequencing is a useful diagnostic tool for complicated forms of hereditary spastic paraplegia. Clin. Genet. 2013;85:154–158. - PubMed
-
- Stevanin G., Santorelli F.M., Azzedine H., Coutinho P., Chomilier J., Denora P.S., Martin E., Ouvrard-Hernandez A.M., Tessa A., Bouslam N., et al. Mutations in SPG11, encoding spatacsin, are a major cause of spastic paraplegia with thin corpus callosum. Nat. Genet. 2007;39:366–372. - PubMed
-
- Hehr U., Bauer P., Winner B., Schule R., Olmez A., Koehler W., Uyanik G., Engel A., Lenz D., Seibel A., et al. Long-term course and mutational spectrum of spatacsin-linked spastic paraplegia. Ann. Neurol. 2007;62:656–665. - PubMed
-
- Winner B., Uyanik G., Gross C., Lange M., Schulte-Mattler W., Schuierer G., Marienhagen J., Hehr U., Winkler J. Clinical progression and genetic analysis in hereditary spastic paraplegia with thin corpus callosum in spastic gait gene 11 (SPG11) Arch. Neurol. 2004;61:117–121. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
