

Trapping of Vibrio cholerae cytolysin in the membrane-bound monomeric state blocks membrane insertion and functional pore formation by the toxin
- PMID: 24794872
- PMCID: PMC4059140
- DOI: 10.1074/jbc.M114.567099
Trapping of Vibrio cholerae cytolysin in the membrane-bound monomeric state blocks membrane insertion and functional pore formation by the toxin
Abstract
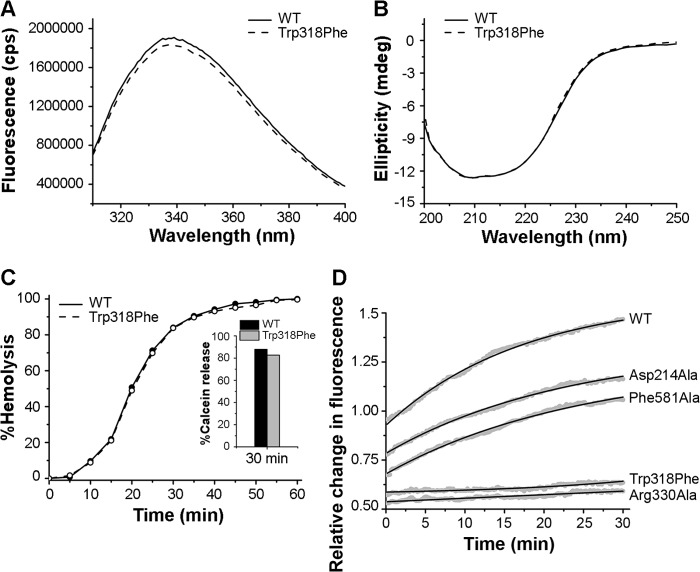
Vibrio cholerae cytolysin (VCC) is a potent membrane-damaging cytolytic toxin that belongs to the family of β barrel pore-forming protein toxins. VCC induces lysis of its target eukaryotic cells by forming transmembrane oligomeric β barrel pores. The mechanism of membrane pore formation by VCC follows the overall scheme of the archetypical β barrel pore-forming protein toxin mode of action, in which the water-soluble monomeric form of the toxin first binds to the target cell membrane, then assembles into a prepore oligomeric intermediate, and finally converts into the functional transmembrane oligomeric β barrel pore. However, there exists a vast knowledge gap in our understanding regarding the intricate details of the membrane pore formation process employed by VCC. In particular, the membrane oligomerization and membrane insertion steps of the process have only been described to a limited extent. In this study, we determined the key residues in VCC that are critical to trigger membrane oligomerization of the toxin. Alteration of such key residues traps the toxin in its membrane-bound monomeric state and abrogates subsequent oligomerization, membrane insertion, and functional transmembrane pore-formation events. The results obtained from our study also suggest that the membrane insertion of VCC depends critically on the oligomerization process and that it cannot be initiated in the membrane-bound monomeric form of the toxin. In sum, our study, for the first time, dissects membrane binding from the subsequent oligomerization and membrane insertion steps and, thus, defines the exact sequence of events in the membrane pore formation process by VCC.
Keywords: Membrane; Membrane Protein; Pore-forming Toxin; Protein Assembly; Protein Structure; Vibrio cholerae Cytolysin; bacterial Toxin.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures




Similar articles
-
Signaling beyond Punching Holes: Modulation of Cellular Responses by Vibrio cholerae Cytolysin.Toxins (Basel). 2015 Aug 21;7(8):3344-58. doi: 10.3390/toxins7083344. Toxins (Basel). 2015. PMID: 26308054 Free PMC article. Review.
-
Glu289 residue in the pore-forming motif of Vibrio cholerae cytolysin is important for efficient β-barrel pore formation.J Biol Chem. 2022 Oct;298(10):102441. doi: 10.1016/j.jbc.2022.102441. Epub 2022 Aug 31. J Biol Chem. 2022. PMID: 36055404 Free PMC article.
-
Physicochemical constraints of elevated pH affect efficient membrane interaction and arrest an abortive membrane-bound oligomeric intermediate of the beta-barrel pore-forming toxin Vibrio cholerae cytolysin.Arch Biochem Biophys. 2015 Oct 1;583:9-17. doi: 10.1016/j.abb.2015.07.019. Epub 2015 Jul 31. Arch Biochem Biophys. 2015. PMID: 26235489
-
Single point mutation in Vibrio cholerae cytolysin compromises the membrane pore-formation mechanism of the toxin.FEBS J. 2012 Nov;279(21):4039-51. doi: 10.1111/j.1742-4658.2012.08809.x. Epub 2012 Sep 27. FEBS J. 2012. PMID: 22934938
-
Vibrio cholerae cytolysin: Multiple facets of the membrane interaction mechanism of a β-barrel pore-forming toxin.IUBMB Life. 2018 Apr;70(4):260-266. doi: 10.1002/iub.1725. Epub 2018 Feb 22. IUBMB Life. 2018. PMID: 29469977 Review.
Cited by
-
Single-particle cryo-EM reveals conformational variability of the oligomeric VCC β-barrel pore in a lipid bilayer.J Cell Biol. 2021 Dec 6;220(12):e202102035. doi: 10.1083/jcb.202102035. Epub 2021 Oct 7. J Cell Biol. 2021. PMID: 34617964 Free PMC article.
-
Signaling beyond Punching Holes: Modulation of Cellular Responses by Vibrio cholerae Cytolysin.Toxins (Basel). 2015 Aug 21;7(8):3344-58. doi: 10.3390/toxins7083344. Toxins (Basel). 2015. PMID: 26308054 Free PMC article. Review.
-
Pore-forming toxins: ancient, but never really out of fashion.Nat Rev Microbiol. 2016 Feb;14(2):77-92. doi: 10.1038/nrmicro.2015.3. Epub 2015 Dec 7. Nat Rev Microbiol. 2016. PMID: 26639780 Review.
-
Structural basis for calcium-stimulating pore formation of Vibrio α-hemolysin.Nat Commun. 2023 Sep 23;14(1):5946. doi: 10.1038/s41467-023-41579-x. Nat Commun. 2023. PMID: 37741869 Free PMC article.
-
Curcumin Inhibits Membrane-Damaging Pore-Forming Function of the β-Barrel Pore-Forming Toxin Vibrio cholerae Cytolysin.Front Microbiol. 2022 Jan 24;12:809782. doi: 10.3389/fmicb.2021.809782. eCollection 2021. Front Microbiol. 2022. PMID: 35140698 Free PMC article.
References
-
- Alouf J. E., Popoff M. R. (2006) The Comprehensive Sourcebook of Bacterial Protein Toxins, 3rd Ed., Academic Press, San Diego, CA
-
- Kaper J. B., Fasano A., Trucksis M. (1994) Toxins of Vibrio cholerae. In Vibrio cholerae and Cholera: Molecular to Global Perspectives (Wachsmuth I. K., Blake P., Olsvik Ø., eds), pp. 145–176, ASM Press, Washington, D. C
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
