

Bile acid signaling in lipid metabolism: metabolomic and lipidomic analysis of lipid and bile acid markers linked to anti-obesity and anti-diabetes in mice
- PMID: 24796972
- PMCID: PMC4219936
- DOI: 10.1016/j.bbalip.2014.04.008
Bile acid signaling in lipid metabolism: metabolomic and lipidomic analysis of lipid and bile acid markers linked to anti-obesity and anti-diabetes in mice
Abstract
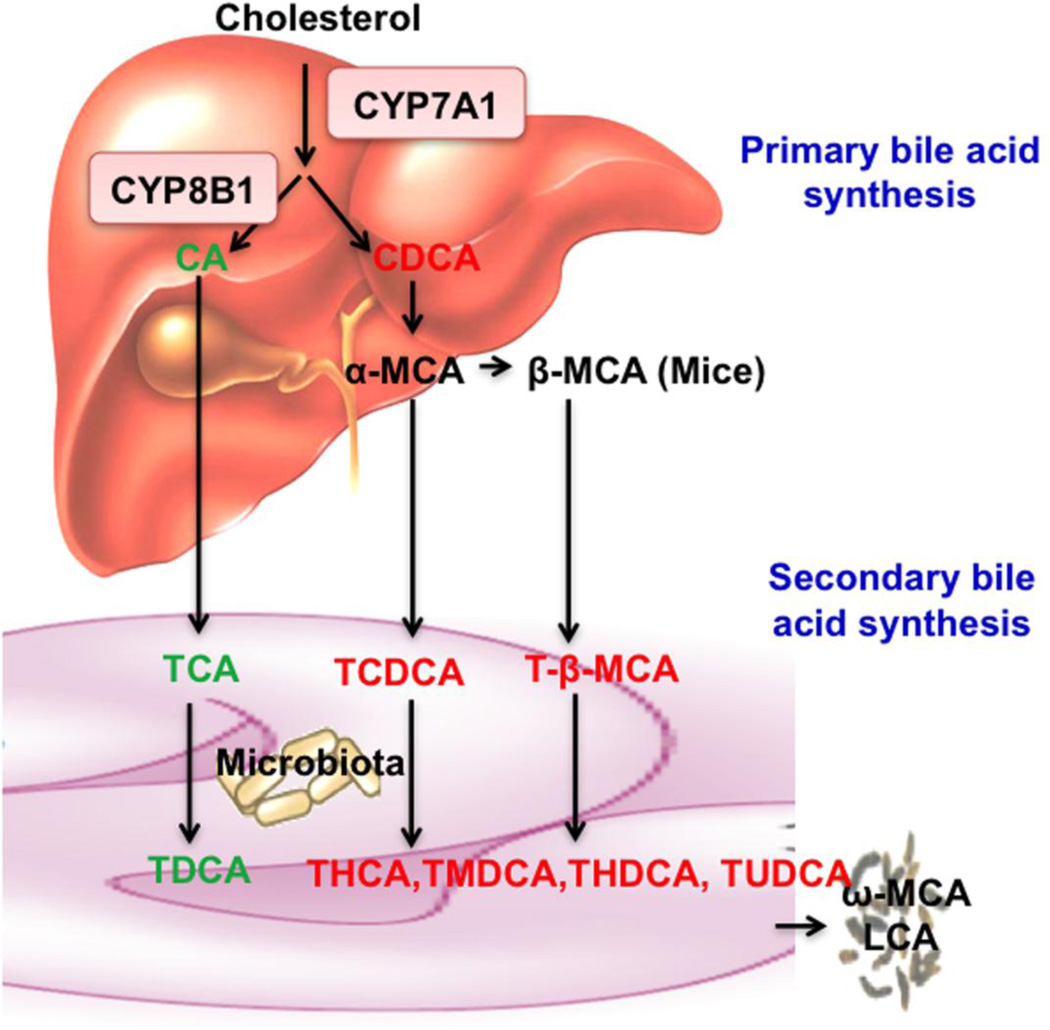
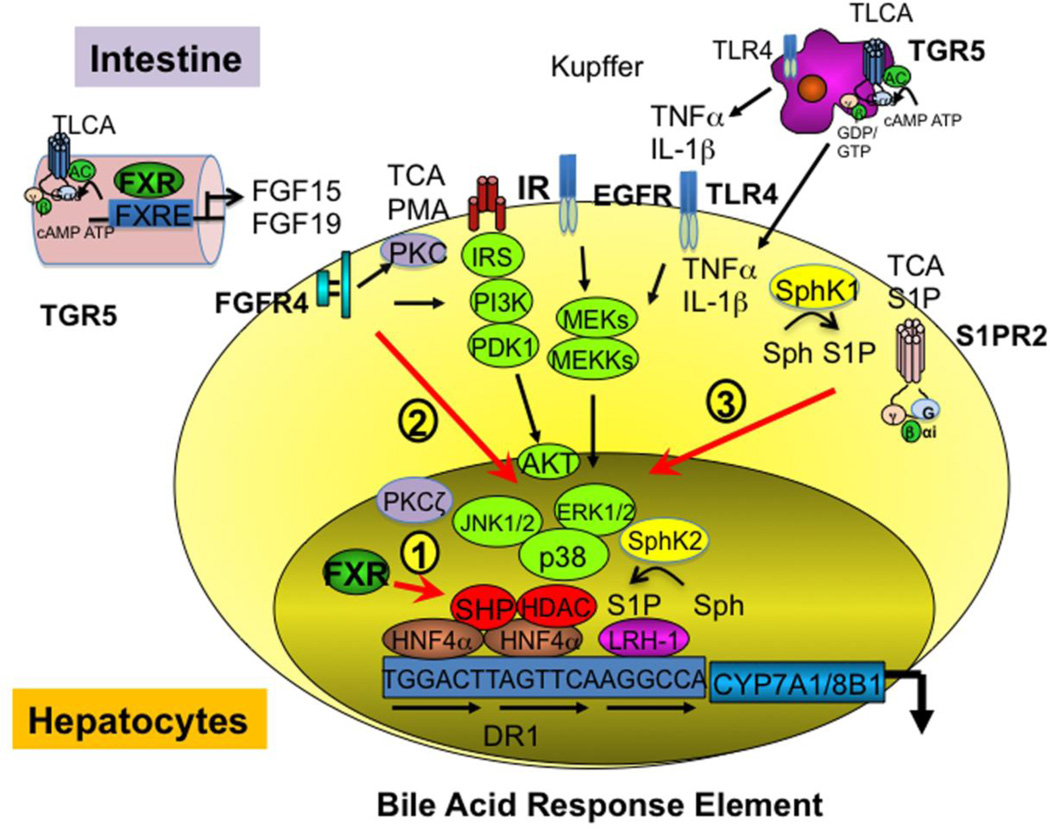
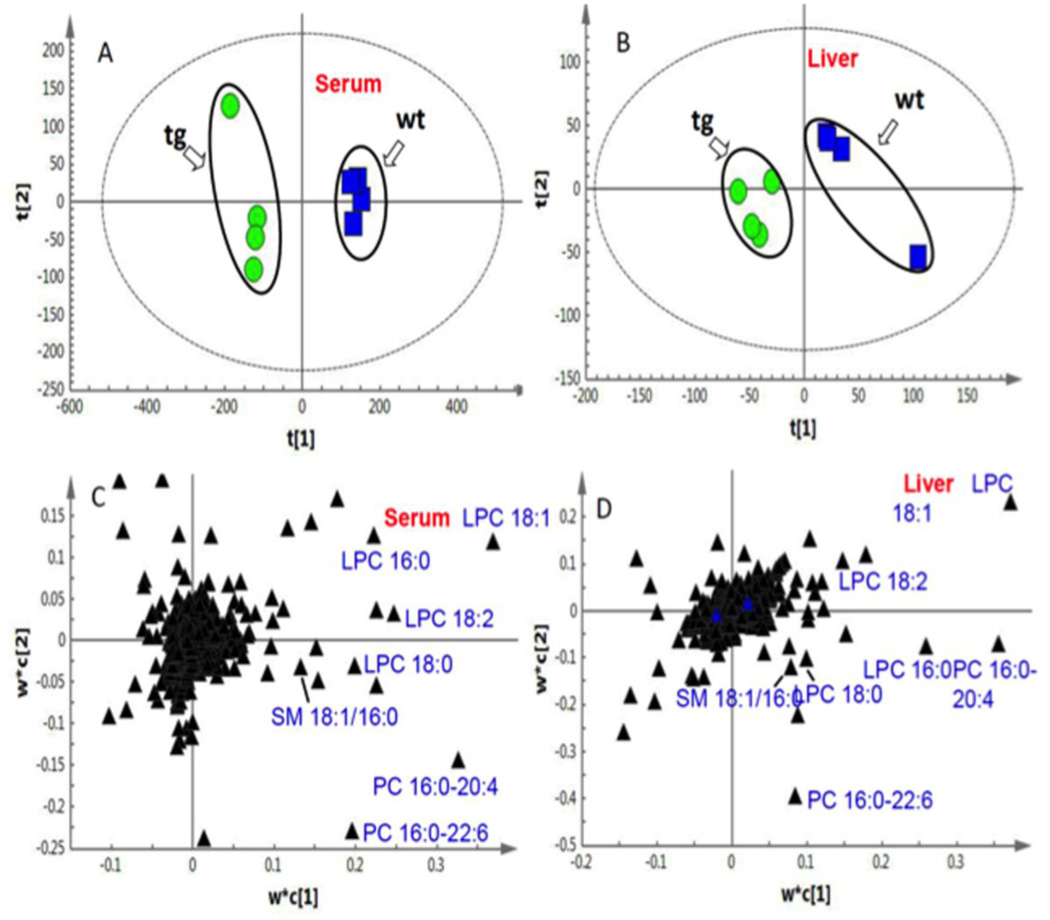
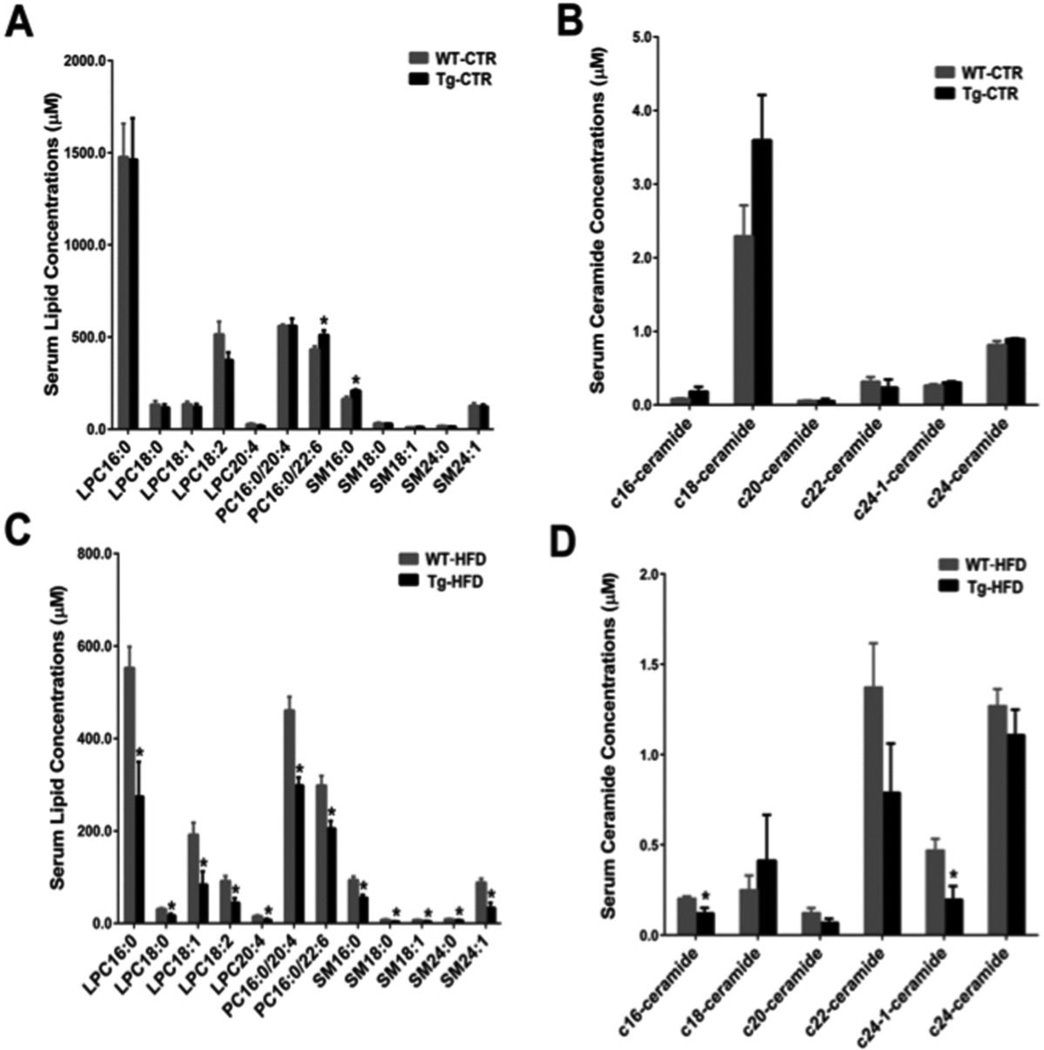
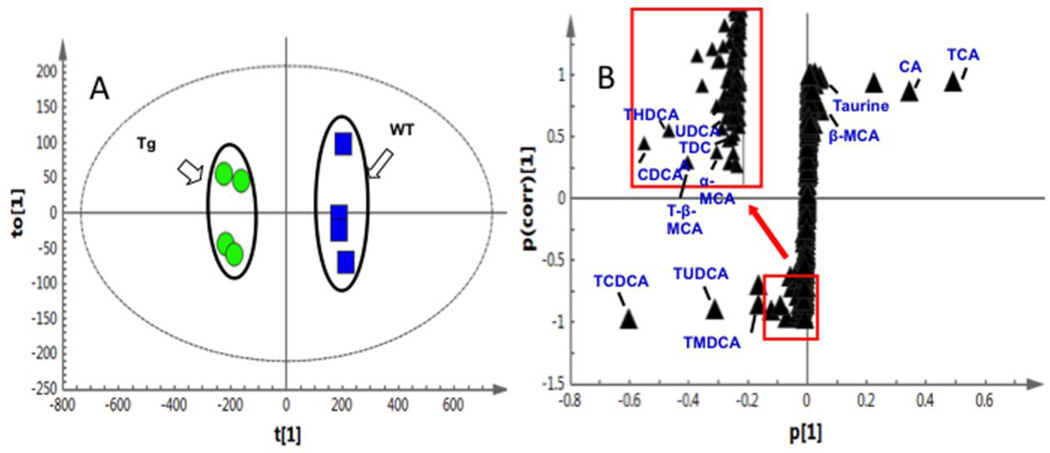
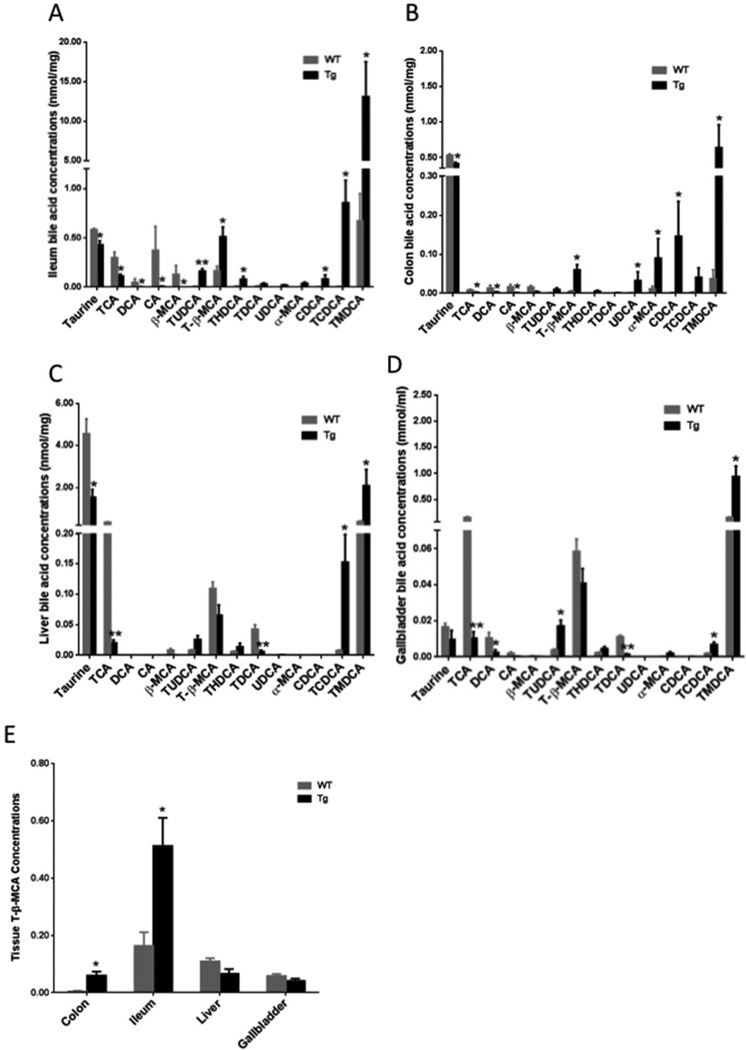
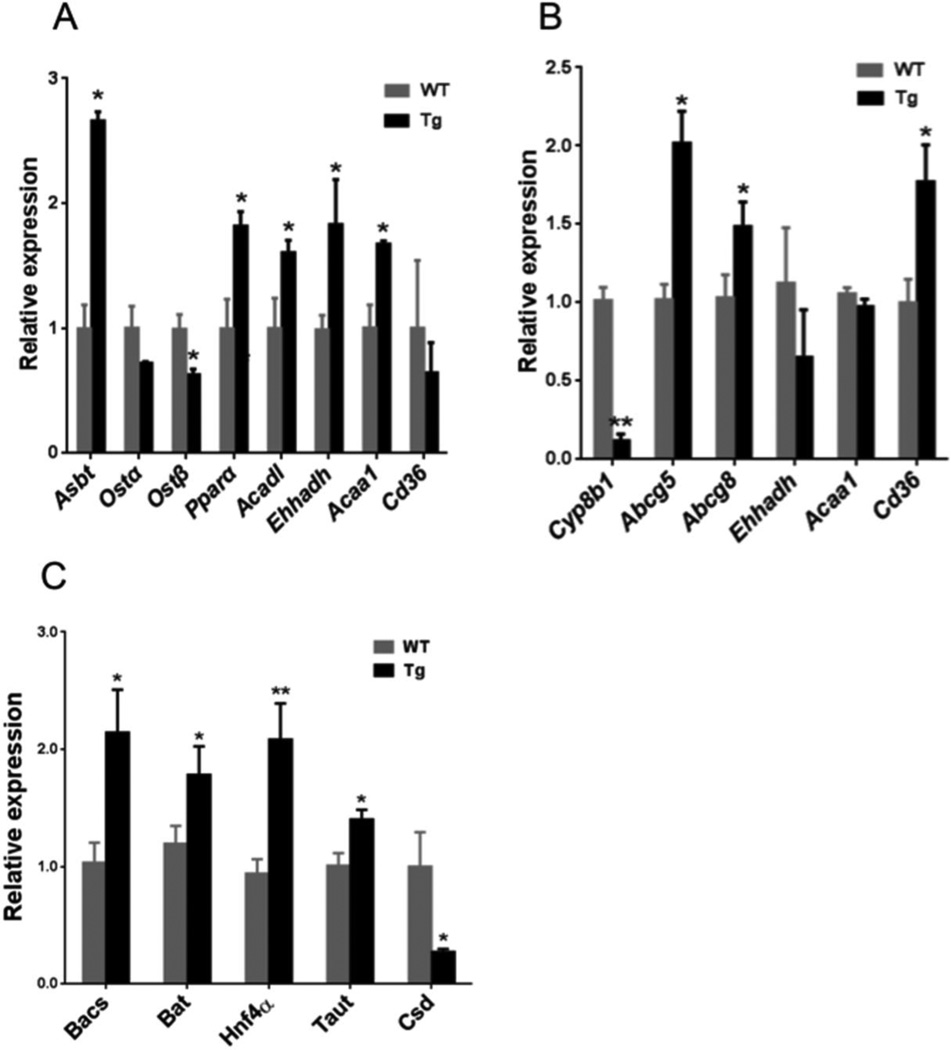
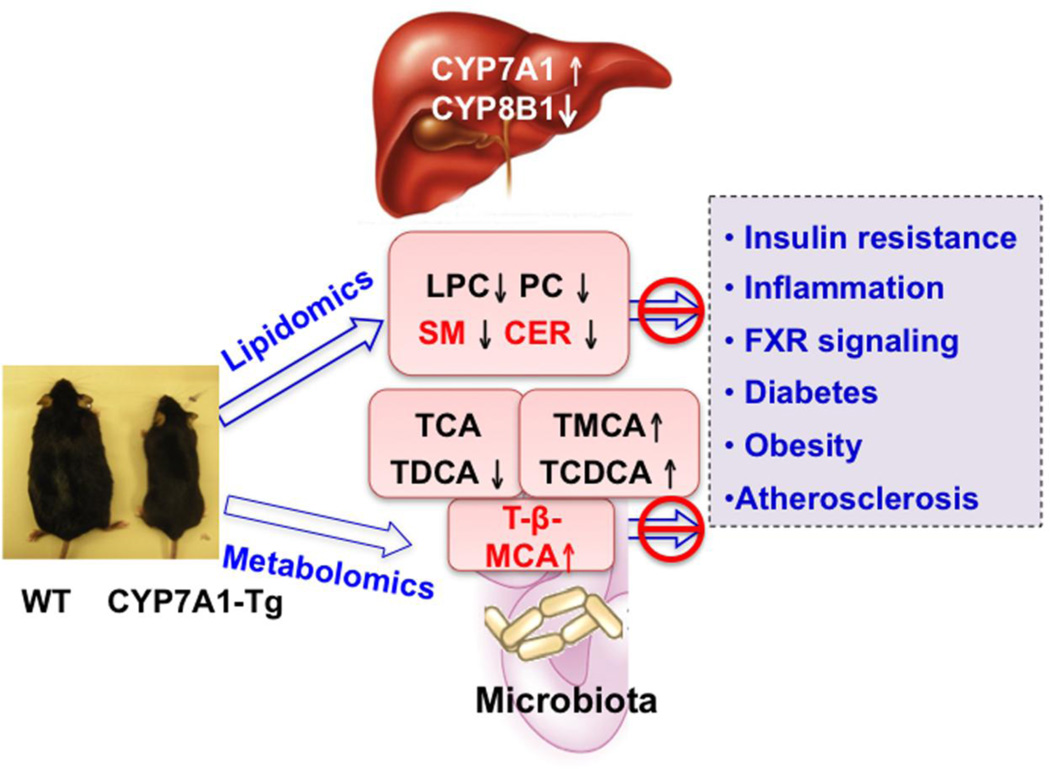
Bile acid synthesis is the major pathway for catabolism of cholesterol. Cholesterol 7α-hydroxylase (CYP7A1) is the rate-limiting enzyme in the bile acid biosynthetic pathway in the liver and plays an important role in regulating lipid, glucose and energy metabolism. Transgenic mice overexpressing CYP7A1 (CYP7A1-tg mice) were resistant to high-fat diet (HFD)-induced obesity, fatty liver, and diabetes. However the mechanism of resistance to HFD-induced obesity of CYP7A1-tg mice has not been determined. In this study, metabolomic and lipidomic profiles of CYP7A1-tg mice were analyzed to explore the metabolic alterations in CYP7A1-tg mice that govern the protection against obesity and insulin resistance by using ultra-performance liquid chromatography-coupled with electrospray ionization quadrupole time-of-flight mass spectrometry combined with multivariate analyses. Lipidomics analysis identified seven lipid markers including lysophosphatidylcholines, phosphatidylcholines, sphingomyelins and ceramides that were significantly decreased in serum of HFD-fed CYP7A1-tg mice. Metabolomics analysis identified 13 metabolites in bile acid synthesis including taurochenodeoxycholic acid, taurodeoxycholic acid, tauroursodeoxycholic acid, taurocholic acid, and tauro-β-muricholic acid (T-β-MCA) that differed between CYP7A1-tg and wild-type mice. Notably, T-β-MCA, an antagonist of the farnesoid X receptor (FXR) was significantly increased in intestine of CYP7A1-tg mice. This study suggests that reducing 12α-hydroxylated bile acids and increasing intestinal T-β-MCA may reduce high fat diet-induced increase of phospholipids, sphingomyelins and ceramides, and ameliorate diabetes and obesity. This article is part of a Special Issue entitled Linking transcription to physiology in lipodomics.
Keywords: CYP7A1; bile acid metabolism; farnesoid X receptor (FXR); lipidomics; tauro-β-muricholic acid.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
-
- Makishima M, Okamoto AY, Repa JJ, Tu H, Learned RM, Luk A, et al. Identification of a nuclear receptor for bile acids. Science. 1999;284:1362–1365. - PubMed
-
- Parks DJ, Blanchard SG, Bledsoe RK, Chandra G, Consler TG, Kliewer SA, et al. Bile acids: Natural ligands for an orphan nuclear receptor. Science. 1999;284:1365–1368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
