

Actin scaffolding by clathrin heavy chain is required for skeletal muscle sarcomere organization
- PMID: 24798732
- PMCID: PMC4018784
- DOI: 10.1083/jcb.201309096
Actin scaffolding by clathrin heavy chain is required for skeletal muscle sarcomere organization
Abstract
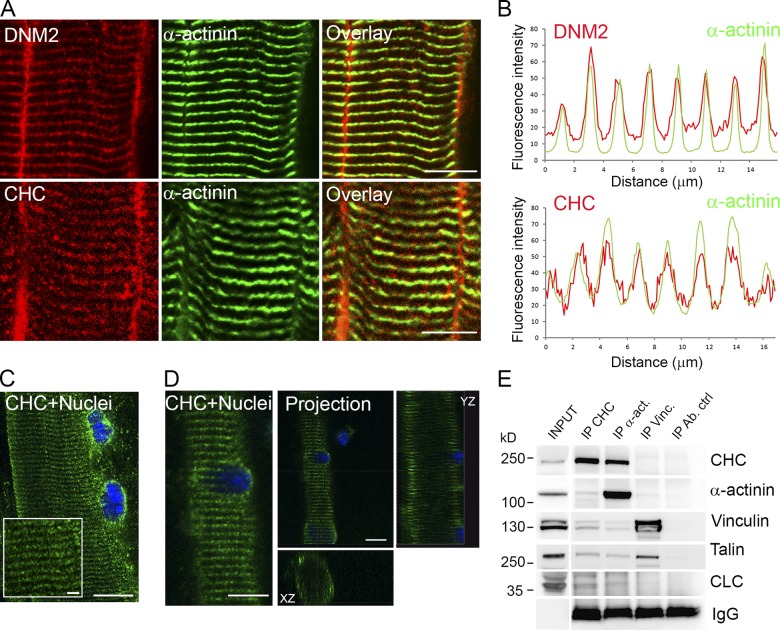
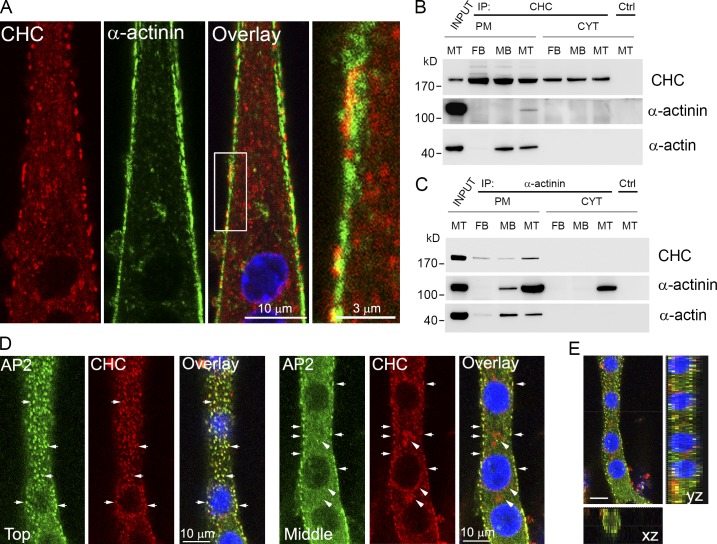
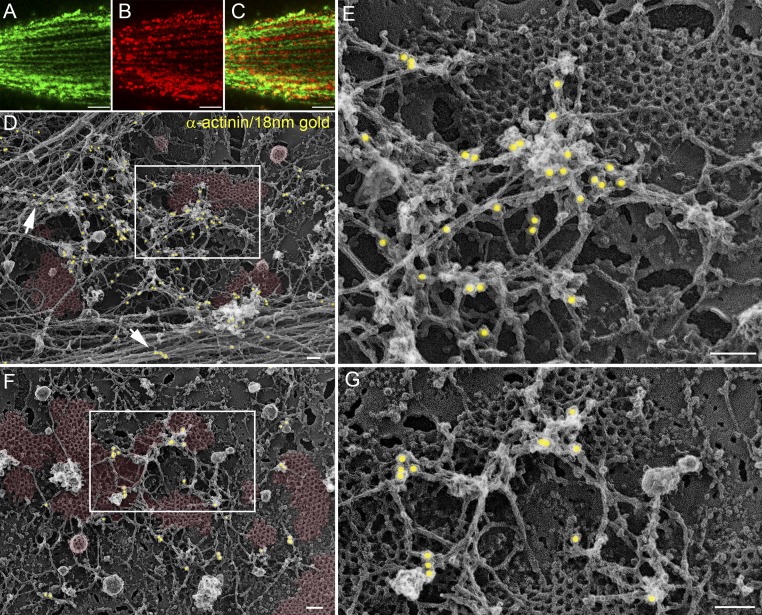
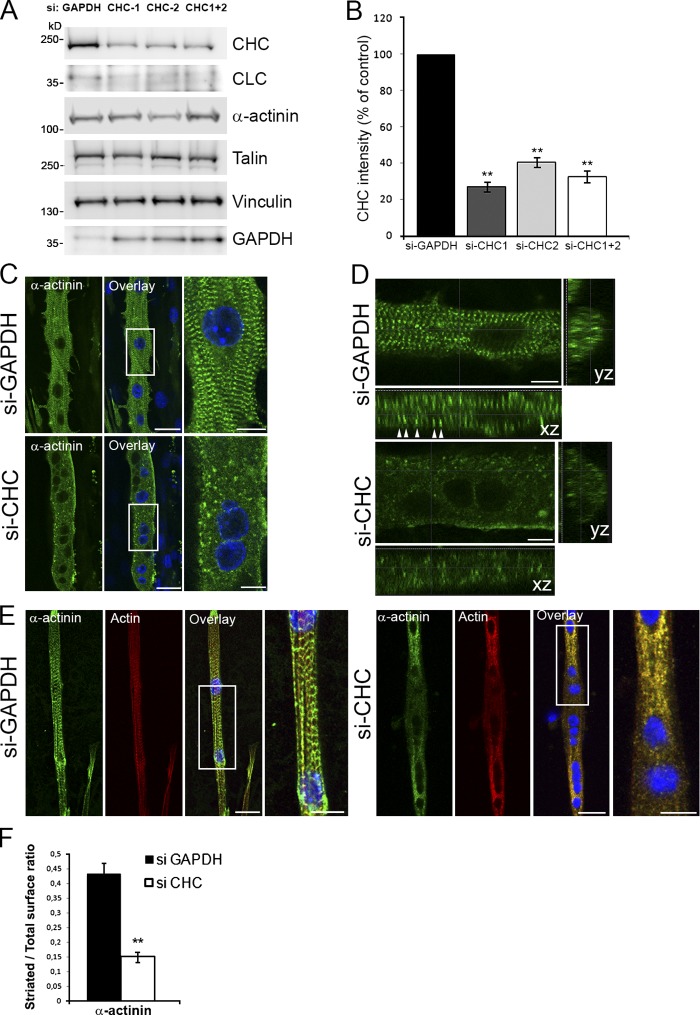
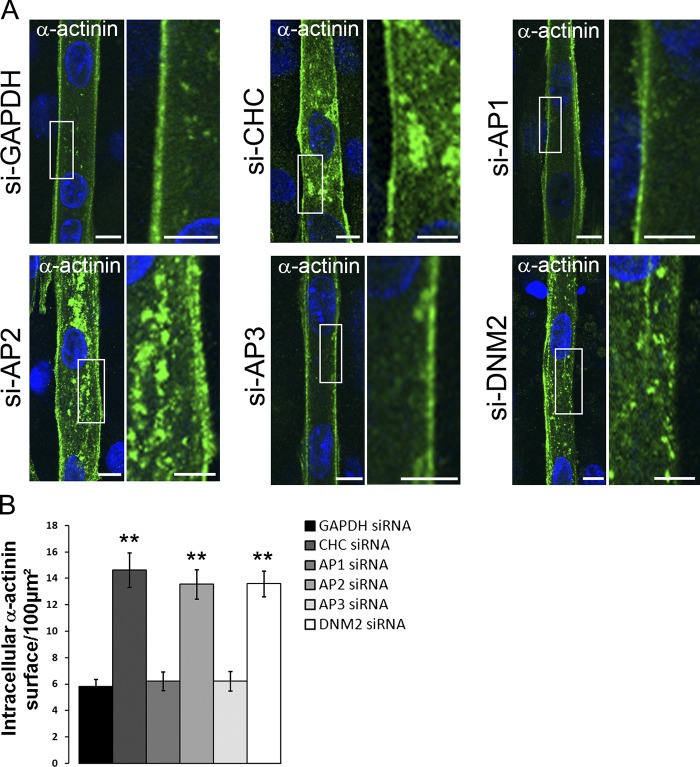
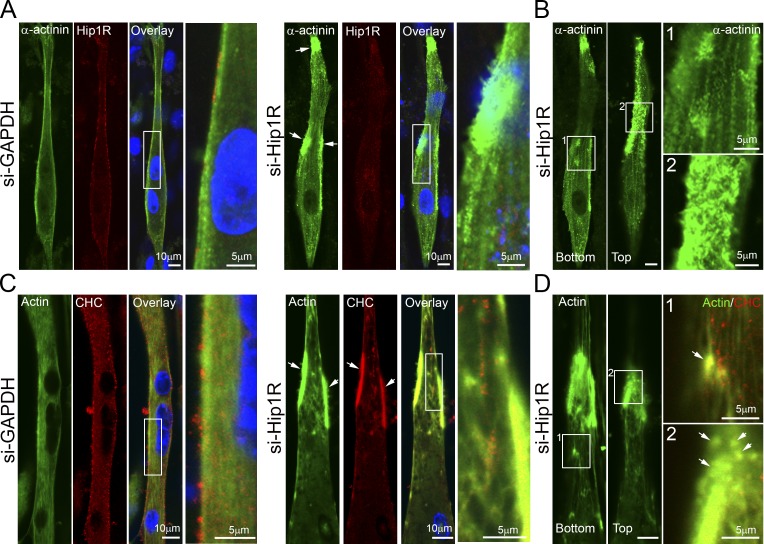
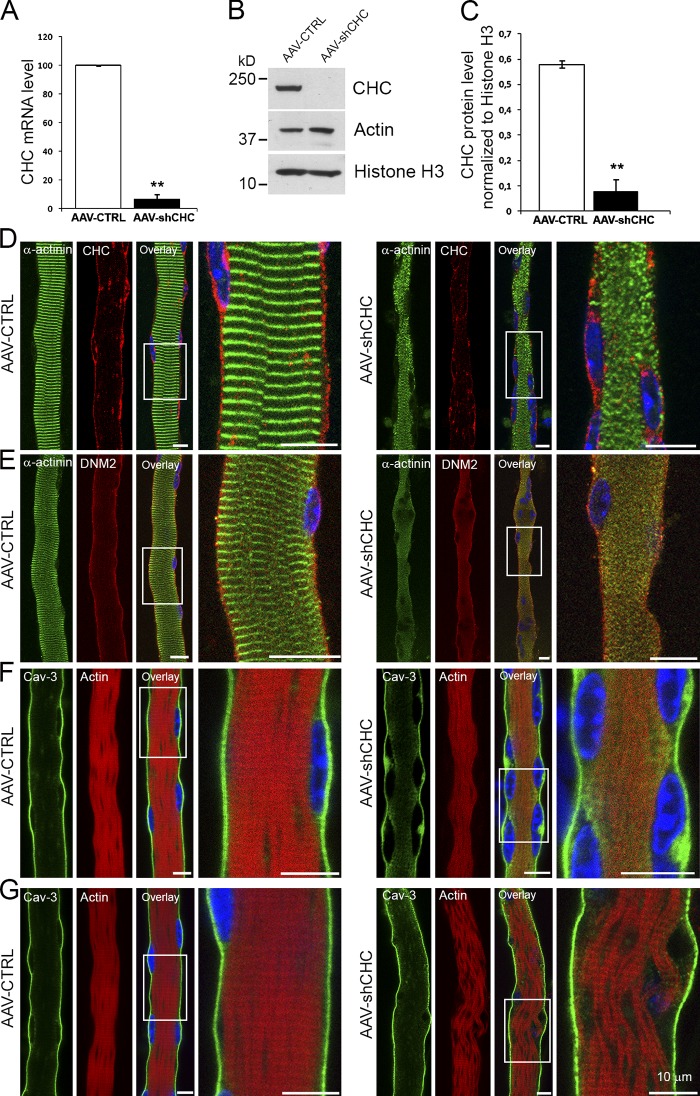
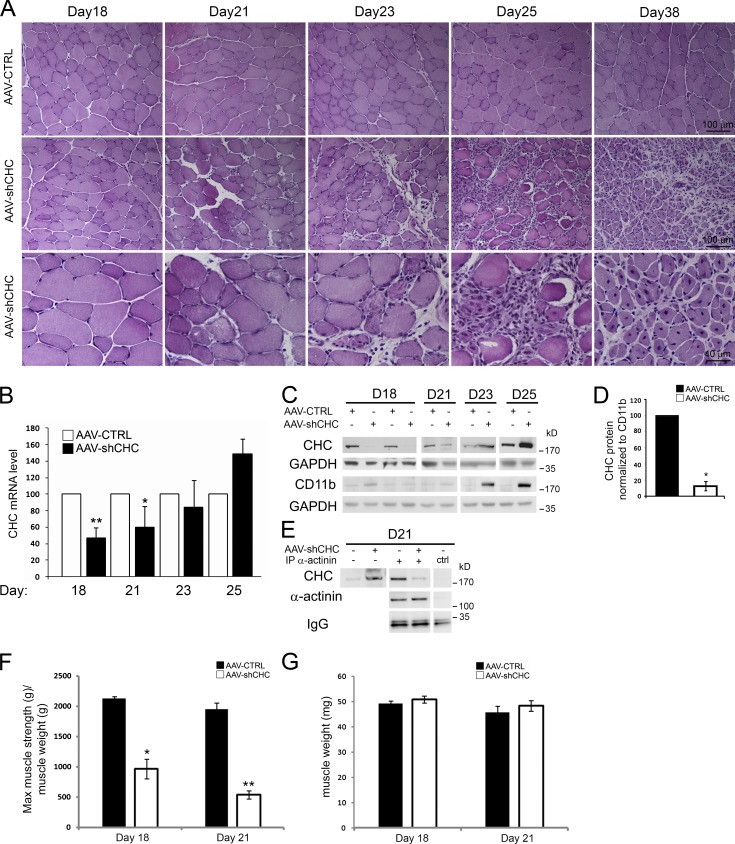
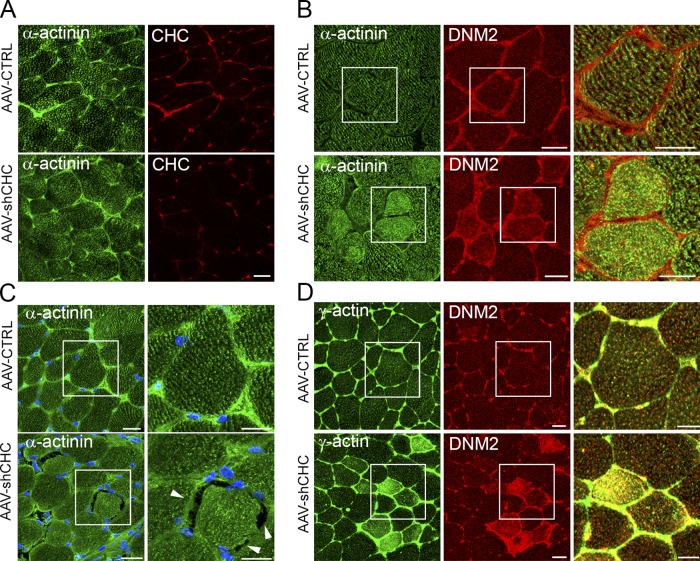
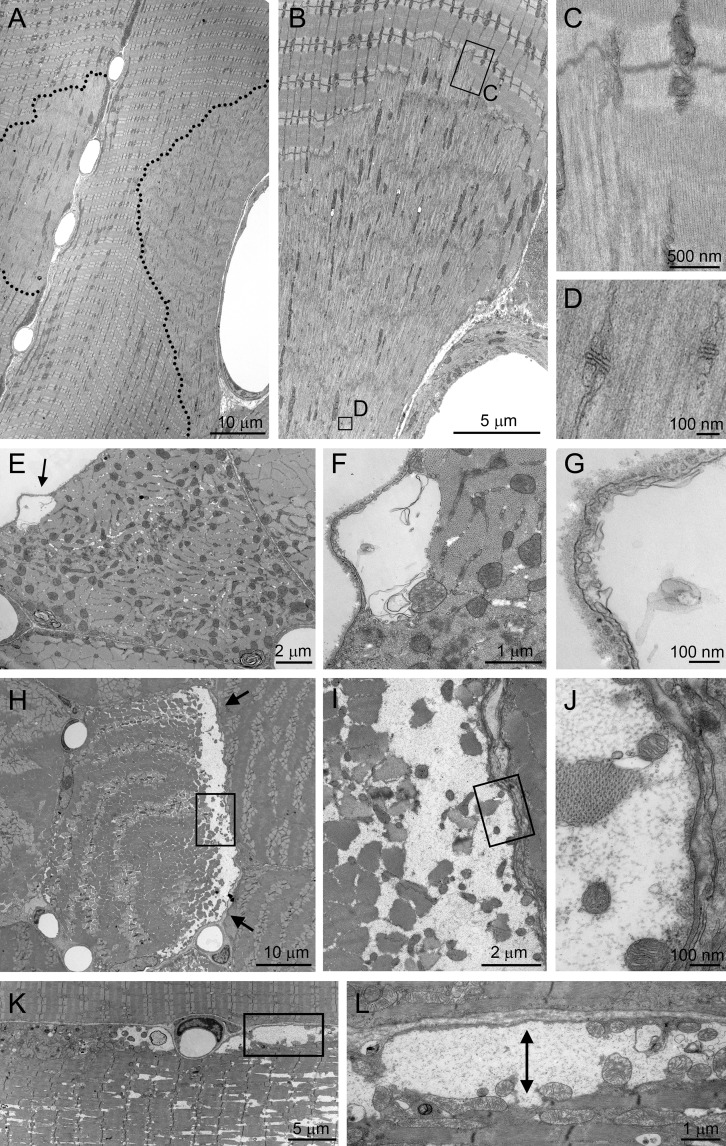
The ubiquitous clathrin heavy chain (CHC), the main component of clathrin-coated vesicles, is well characterized for its role in intracellular membrane traffic and endocytosis from the plasma membrane (PM). Here, we demonstrate that in skeletal muscle CHC regulates the formation and maintenance of PM-sarcomere attachment sites also known as costameres. We show that clathrin forms large coated lattices associated with actin filaments and the muscle-specific isoform of α-actinin at the PM of differentiated myotubes. Depletion of CHC in myotubes induced a loss of actin and α-actinin sarcomeric organization, whereas CHC depletion in vivo induced a loss of contractile force due to the detachment of sarcomeres from the PM. Our results suggest that CHC contributes to the formation and maintenance of the contractile apparatus through interactions with costameric proteins and highlight an unconventional role for CHC in skeletal muscle that may be relevant to pathophysiology of neuromuscular disorders.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
