

Sympathetic stimulation of thiazide-sensitive sodium chloride cotransport in the generation of salt-sensitive hypertension
- PMID: 24799612
- PMCID: PMC4061471
- DOI: 10.1161/HYPERTENSIONAHA.114.03335
Sympathetic stimulation of thiazide-sensitive sodium chloride cotransport in the generation of salt-sensitive hypertension
Abstract
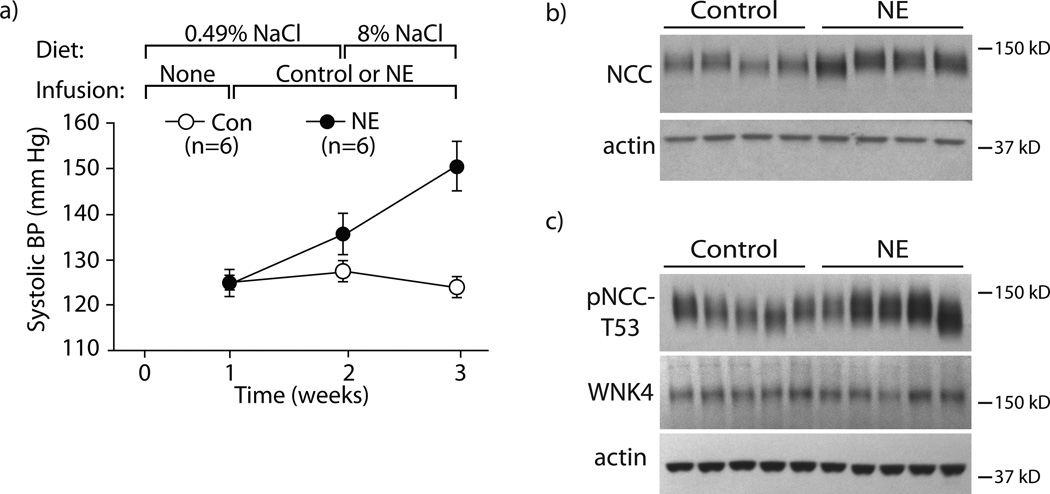
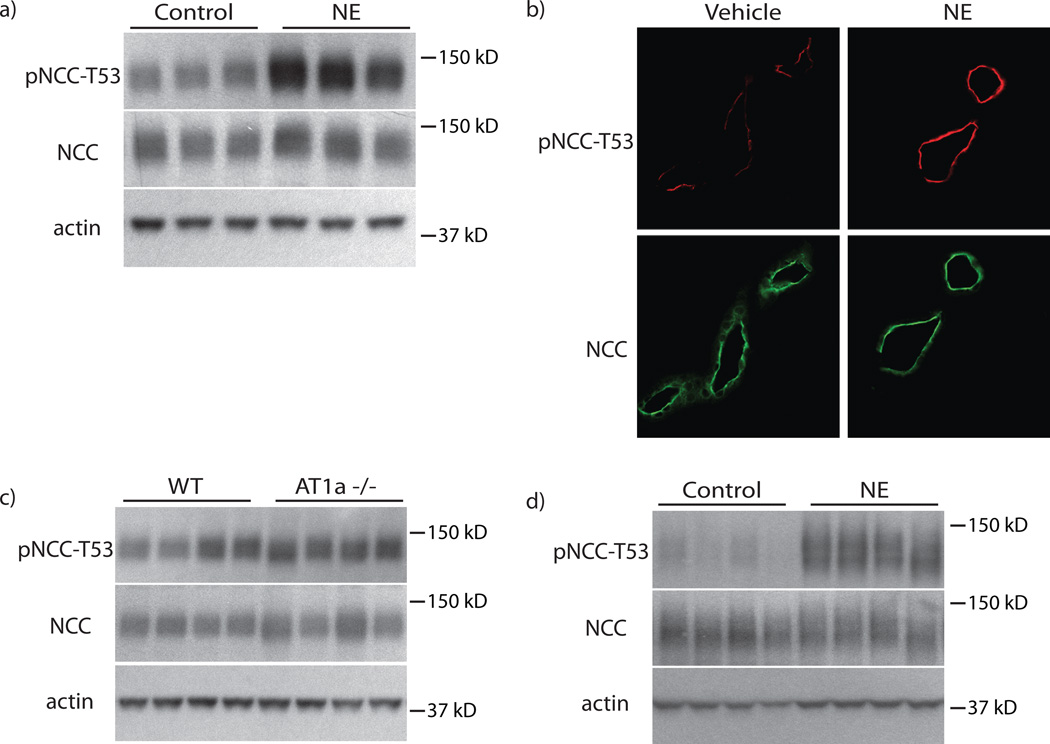
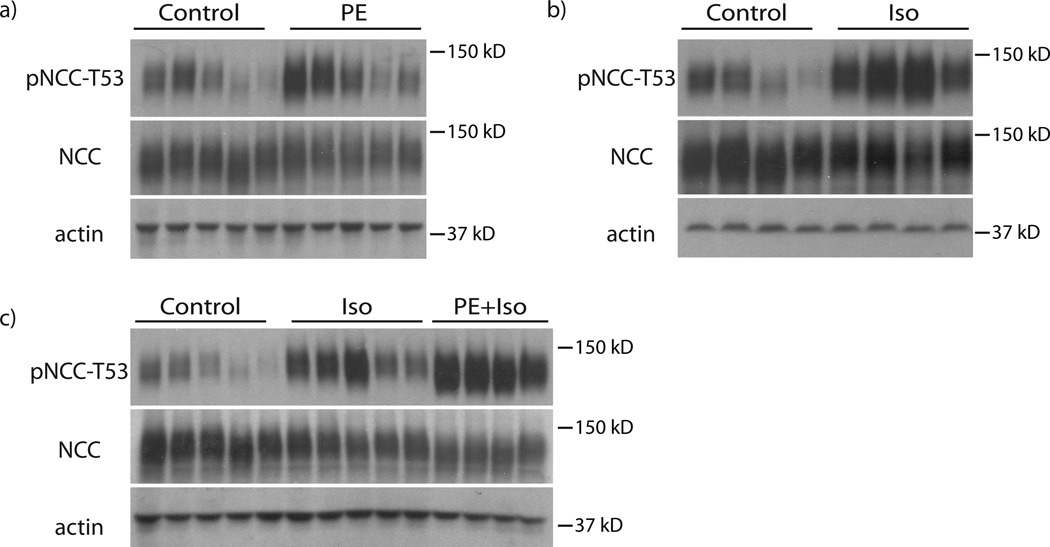
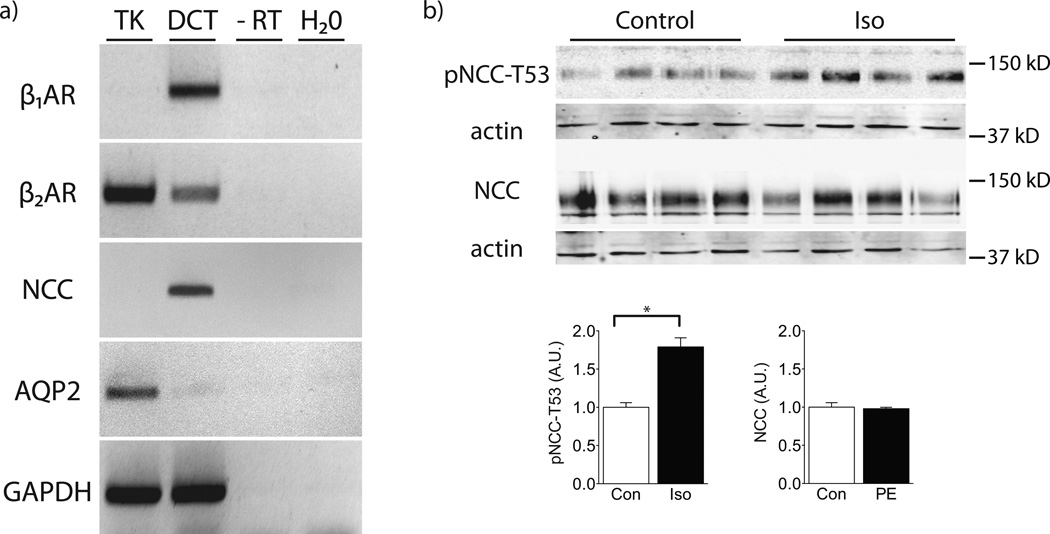
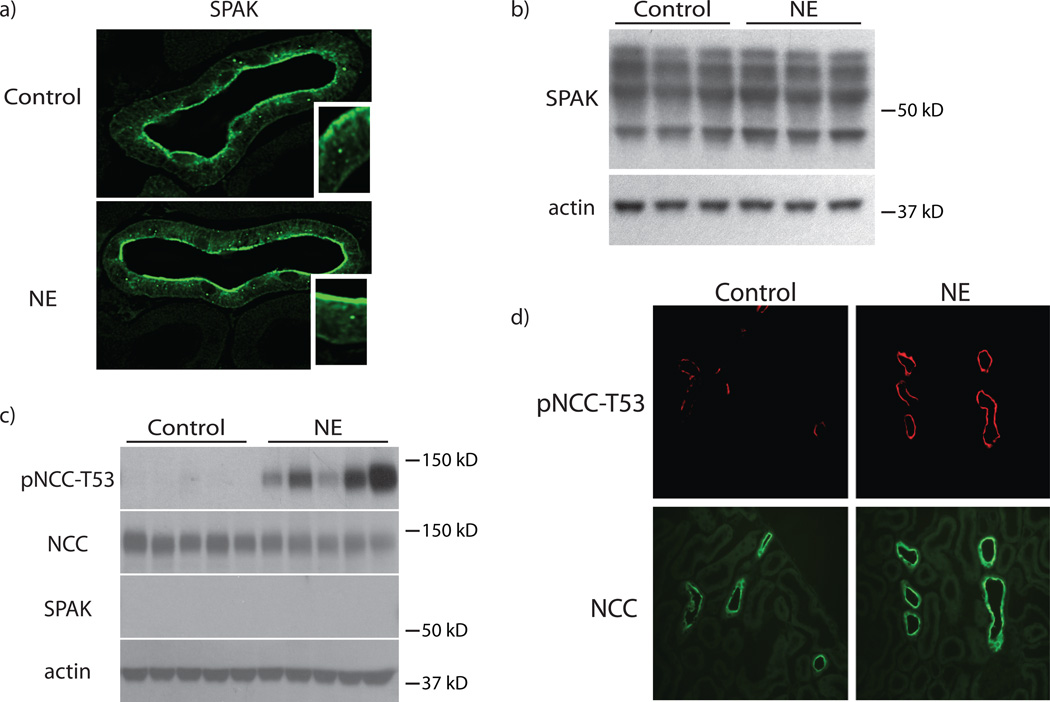
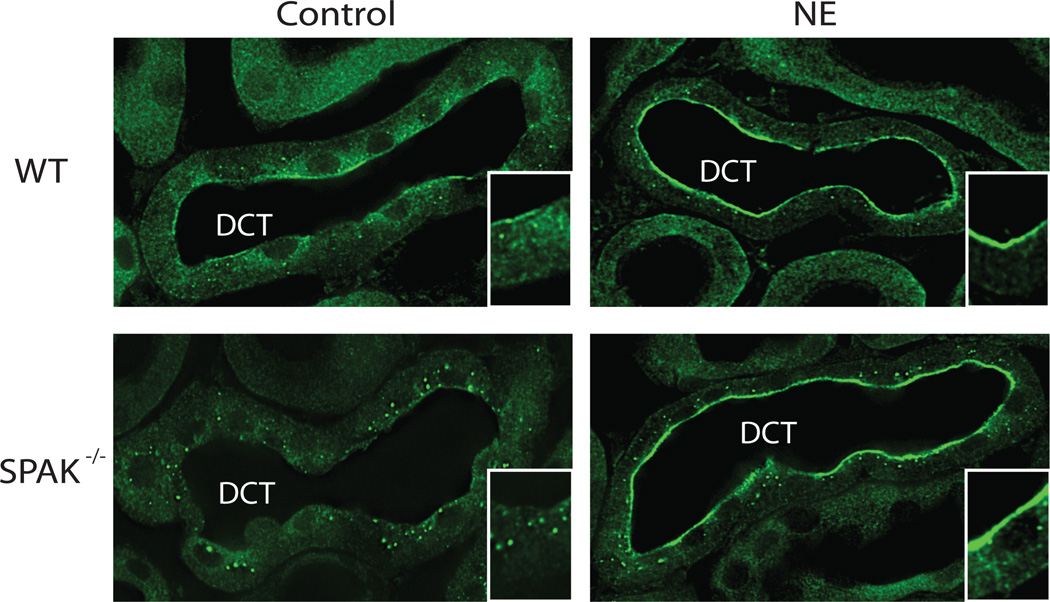
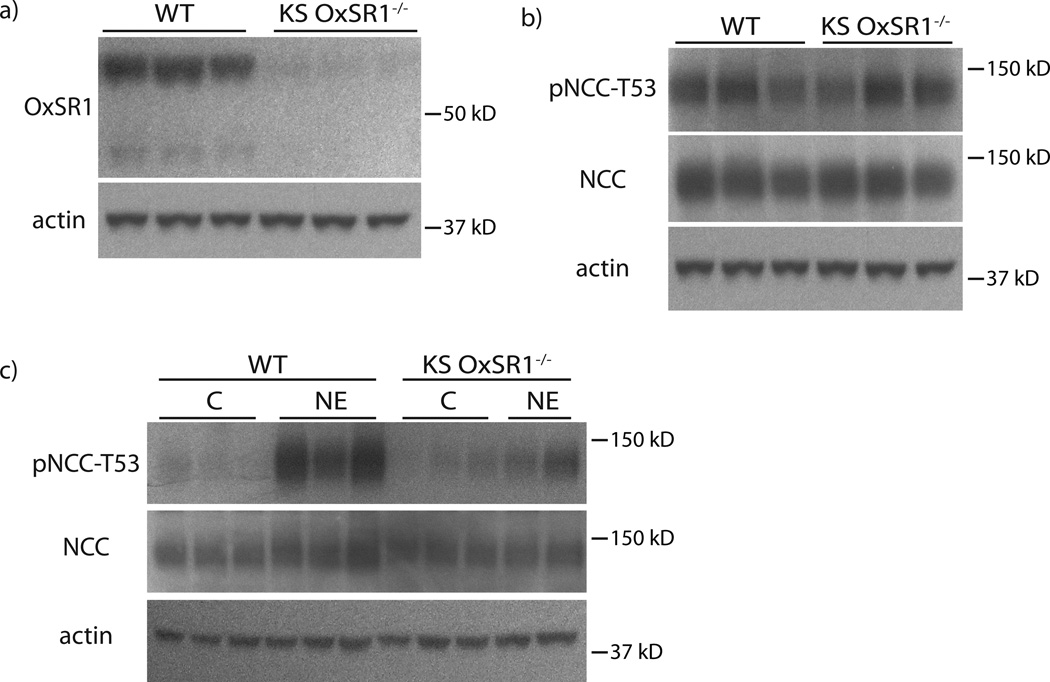
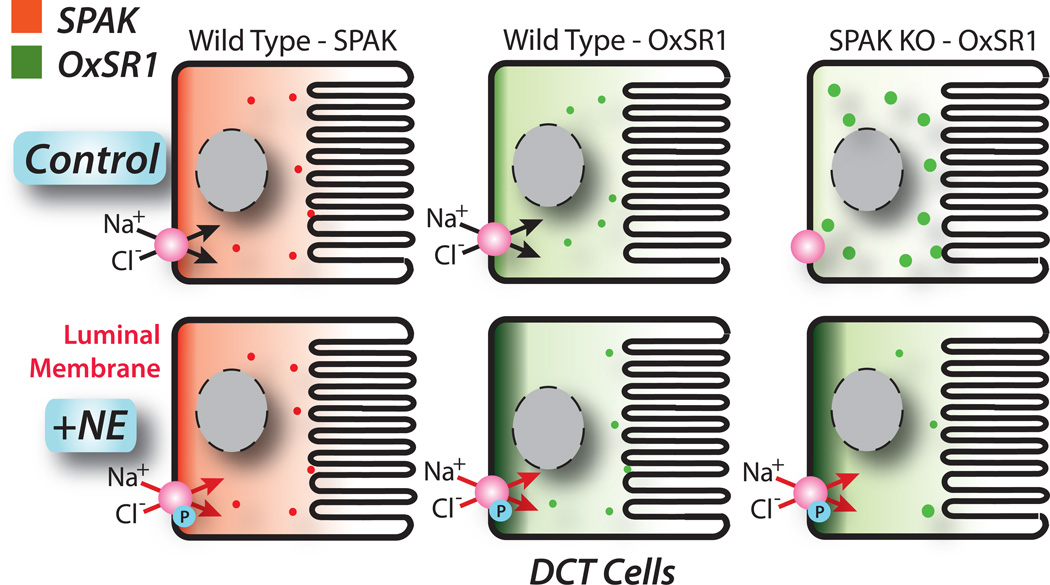
Excessive renal efferent sympathetic nerve activity contributes to hypertension in many circumstances. Although both hemodynamic and tubular effects likely participate, most evidence supports a major role for α-adrenergic receptors in mediating the direct epithelial stimulation of sodium retention. Recently, it was reported, however, that norepinephrine activates the thiazide-sensitive NaCl cotransporter (NCC) by stimulating β-adrenergic receptors. Here, we confirmed this effect and developed an acute adrenergic stimulation model to study the signaling cascade. The results show that norepinephrine increases the abundance of phosphorylated NCC rapidly (161% increase), an effect largely dependent on β-adrenergic receptors. This effect is not mediated by the activation of angiotensin II receptors. We used immunodissected mouse distal convoluted tubule to show that distal convoluted tubule cells are especially enriched for β₁-adrenergic receptors, and that the effects of adrenergic stimulation can occur ex vivo (79% increase), suggesting they are direct. Because the 2 protein kinases, STE20p-related proline- and alanine-rich kinase (encoded by STK39) and oxidative stress-response kinase 1, phosphorylate and activate NCC, we examined their roles in norepinephrine effects. Surprisingly, norepinephrine did not affect STE20p-related proline- and alanine-rich kinase abundance or its localization in the distal convoluted tubule; instead, we observed a striking activation of oxidative stress-response kinase 1. We confirmed that STE20p-related proline- and alanine-rich kinase is not required for NCC activation, using STK39 knockout mice. Together, the data provide strong support for a signaling system involving β₁-receptors in the distal convoluted tubule that activates NCC, at least in part via oxidative stress-response kinase 1. The results have implications about device- and drug-based treatment of hypertension.
Keywords: diuretics; hypertension; ion transport; sodium-potassium-chloride symporters; sympathetic nervous system.
© 2014 American Heart Association, Inc.
Conflict of interest statement
None.
Figures
Similar articles
-
Norepinephrine-Induced Stimulation of Kir4.1/Kir5.1 Is Required for the Activation of NaCl Transporter in Distal Convoluted Tubule.Hypertension. 2019 Jan;73(1):112-120. doi: 10.1161/HYPERTENSIONAHA.118.11621. Hypertension. 2019. PMID: 30571558 Free PMC article.
-
Activation of the kidney sodium chloride cotransporter by the β2-adrenergic receptor agonist salbutamol increases blood pressure.Kidney Int. 2021 Aug;100(2):321-335. doi: 10.1016/j.kint.2021.04.021. Epub 2021 Apr 30. Kidney Int. 2021. PMID: 33940111
-
SPAK and OSR1 play essential roles in potassium homeostasis through actions on the distal convoluted tubule.J Physiol. 2016 Sep 1;594(17):4945-66. doi: 10.1113/JP272311. Epub 2016 May 29. J Physiol. 2016. PMID: 27068441 Free PMC article.
-
Mini-review: regulation of the renal NaCl cotransporter by hormones.Am J Physiol Renal Physiol. 2016 Jan 1;310(1):F10-4. doi: 10.1152/ajprenal.00354.2015. Epub 2015 Oct 28. Am J Physiol Renal Physiol. 2016. PMID: 26511649 Review.
-
Regulation of the WNK4-SPAK-NCC pathway by the calcium-sensing receptor.Curr Opin Nephrol Hypertens. 2023 Sep 1;32(5):451-457. doi: 10.1097/MNH.0000000000000915. Epub 2023 Jul 7. Curr Opin Nephrol Hypertens. 2023. PMID: 37530086 Review.
Cited by
-
Renal tubular NHE3 is required in the maintenance of water and sodium chloride homeostasis.Kidney Int. 2017 Aug;92(2):397-414. doi: 10.1016/j.kint.2017.02.001. Epub 2017 Apr 3. Kidney Int. 2017. PMID: 28385297 Free PMC article.
-
New insights into sodium transport regulation in the distal nephron: Role of G-protein coupled receptors.World J Biol Chem. 2016 Feb 26;7(1):44-63. doi: 10.4331/wjbc.v7.i1.44. World J Biol Chem. 2016. PMID: 26981195 Free PMC article. Review.
-
The Evaluation and Therapeutic Management of Hypertension in the Transplant Patient.Curr Cardiol Rep. 2015 Nov;17(11):95. doi: 10.1007/s11886-015-0647-z. Curr Cardiol Rep. 2015. PMID: 26362301 Review.
-
Landscape of GPCR expression along the mouse nephron.Am J Physiol Renal Physiol. 2021 Jul 1;321(1):F50-F68. doi: 10.1152/ajprenal.00077.2021. Epub 2021 May 24. Am J Physiol Renal Physiol. 2021. PMID: 34029142 Free PMC article.
-
Navigating the multifaceted intricacies of the Na+-Cl- cotransporter, a highly regulated key effector in the control of hydromineral homeostasis.Physiol Rev. 2024 Jul 1;104(3):1147-1204. doi: 10.1152/physrev.00027.2023. Epub 2024 Feb 8. Physiol Rev. 2024. PMID: 38329422 Free PMC article. Review.
References
-
- DiBona GF. Physiology in perspective: The Wisdom of the Body. Neural control of the kidney. Am J Physiol Regul Integr Comp Physiol. 2005;289:R633–R641. - PubMed
-
- Lim SS, Vos T, Flaxman AD, Danaei G, Shibuya K, Adair-Rohani H, Amann M, Anderson HR, Andrews KG, Aryee M, Atkinson C, Bacchus LJ, Bahalim AN, Balakrishnan K, Balmes J, Barker-Collo S, Baxter A, Bell ML, Blore JD, Blyth F, Bonner C, Borges G, Bourne R, Boussinesq M, Brauer M, Brooks P, Bruce NG, Brunekreef B, Bryan-Hancock C, Bucello C, Buchbinder R, Bull F, Burnett RT, Byers TE, Calabria B, Carapetis J, Carnahan E, Chafe Z, Charlson F, Chen H, Chen JS, Cheng AT, Child JC, Cohen A, Colson KE, Cowie BC, Darby S, Darling S, Davis A, Degenhardt L, Dentener F, Des Jarlais DC, Devries K, Dherani M, Ding EL, Dorsey ER, Driscoll T, Edmond K, Ali SE, Engell RE, Erwin PJ, Fahimi S, Falder G, Farzadfar F, Ferrari A, Finucane MM, Flaxman S, Fowkes FG, Freedman G, Freeman MK, Gakidou E, Ghosh S, Giovannucci E, Gmel G, Graham K, Grainger R, Grant B, Gunnell D, Gutierrez HR, Hall W, Hoek HW, Hogan A, Hosgood HD, 3rd, Hoy D, Hu H, Hubbell BJ, Hutchings SJ, Ibeanusi SE, Jacklyn GL, Jasrasaria R, Jonas JB, Kan H, Kanis JA, Kassebaum N, Kawakami N, Khang YH, Khatibzadeh S, Khoo JP, Kok C, Laden F, Lalloo R, Lan Q, Lathlean T, Leasher JL, Leigh J, Li Y, Lin JK, Lipshultz SE, London S, Lozano R, Lu Y, Mak J, Malekzadeh R, Mallinger L, Marcenes W, March L, Marks R, Martin R, McGale P, McGrath J, Mehta S, Mensah GA, Merriman TR, Micha R, Michaud C, Mishra V, Hanafiah KM, Mokdad AA, Morawska L, Mozaffarian D, Murphy T, Naghavi M, Neal B, Nelson PK, Nolla JM, Norman R, Olives C, Omer SB, Orchard J, Osborne R, Ostro B, Page A, Pandey KD, Parry CD, Passmore E, Patra J, Pearce N, Pelizzari PM, Petzold M, Phillips MR, Pope D, Pope CA, 3rd, Powles J, Rao M, Razavi H, Rehfuess EA, Rehm JT, Ritz B, Rivara FP, Roberts T, Robinson C, Rodriguez-Portales JA, Romieu I, Room R, Rosenfeld LC, Roy A, Rushton L, Salomon JA, Sampson U, Sanchez-Riera L, Sanman E, Sapkota A, Seedat S, Shi P, Shield K, Shivakoti R, Singh GM, Sleet DA, Smith E, Smith KR, Stapelberg NJ, Steenland K, Stockl H, Stovner LJ, Straif K, Straney L, Thurston GD, Tran JH, Van Dingenen R, van Donkelaar A, Veerman JL, Vijayakumar L, Weintraub R, Weissman MM, White RA, Whiteford H, Wiersma ST, Wilkinson JD, Williams HC, Williams W, Wilson N, Woolf AD, Yip P, Zielinski JM, Lopez AD, Murray CJ, Ezzati M, AlMazroa MA, Memish ZA. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012;380:2224–2260. - PMC - PubMed
-
- Jacob F, Clark LA, Guzman PA, Osborn JW. Role of renal nerves in development of hypertension in DOCA-salt model in rats: a telemetric approach. American Journal of Physiology - Heart & Circulatory Physiology. 2005;289:H1519–H1529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
