

Inhibition of CaV3.2 T-type calcium channels in peripheral sensory neurons contributes to analgesic properties of epipregnanolone
- PMID: 24800894
- PMCID: PMC4135044
- DOI: 10.1007/s00213-014-3588-0
Inhibition of CaV3.2 T-type calcium channels in peripheral sensory neurons contributes to analgesic properties of epipregnanolone
Abstract
Rationale: T-type calcium channels (T-channels) play an important role in controlling excitability of nociceptors. We have previously shown that a synthetic series of 5β-reduced steroids induce a voltage-dependent blockade of T-currents in rat dorsal root ganglia (DRG) cells in vitro and induce potent analgesia to thermal stimuli in rats in vivo (Mol Pharmacol 66:1223-1235, 2004).
Objectives: Here, we investigated the effects of the endogenous 5β-reduced neuroactive steroid molecule, epipregnanolone [(3β,5β)-3-hydroxypregnan-20-one], on peripheral nociception.
Methods: We used acutely dissociated DRG cells in vitro from adult rats as well as in vivo pain studies in mice and rats to investigate the effects of epipregnanolone on DRG T-channels.
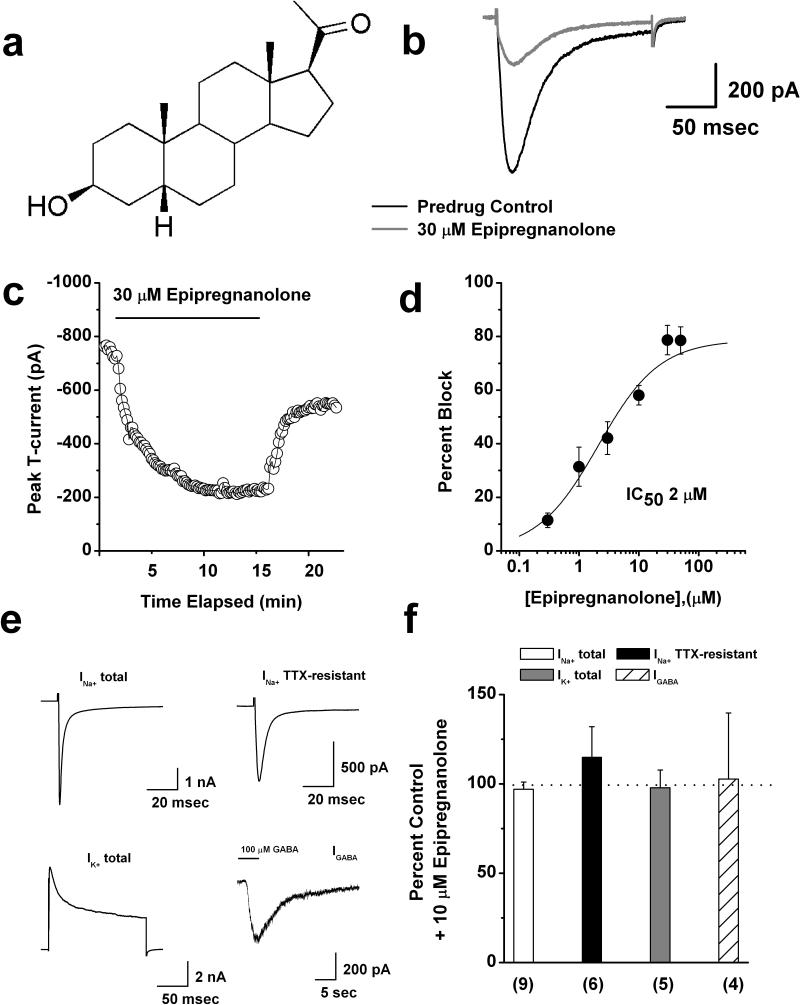
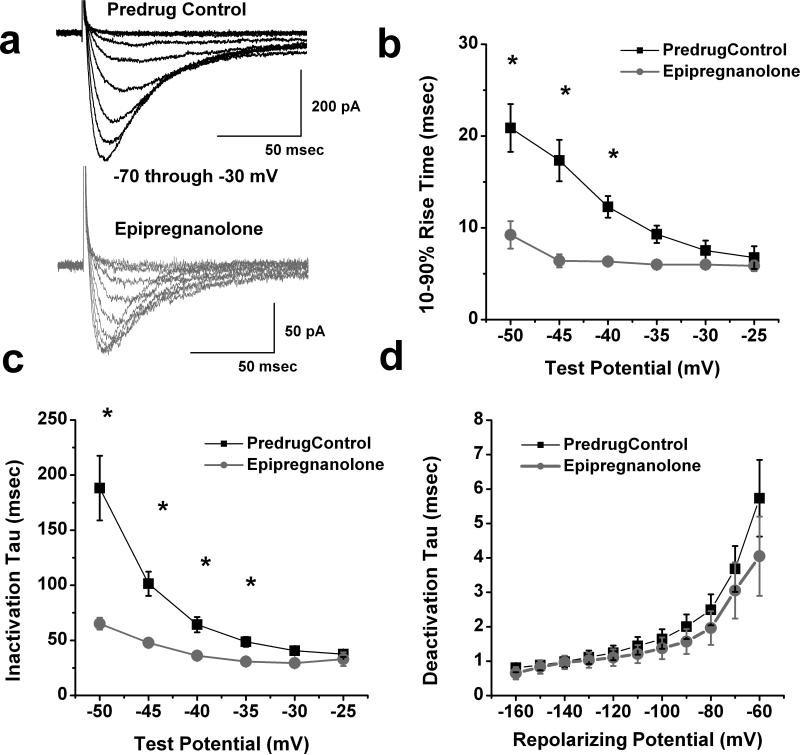
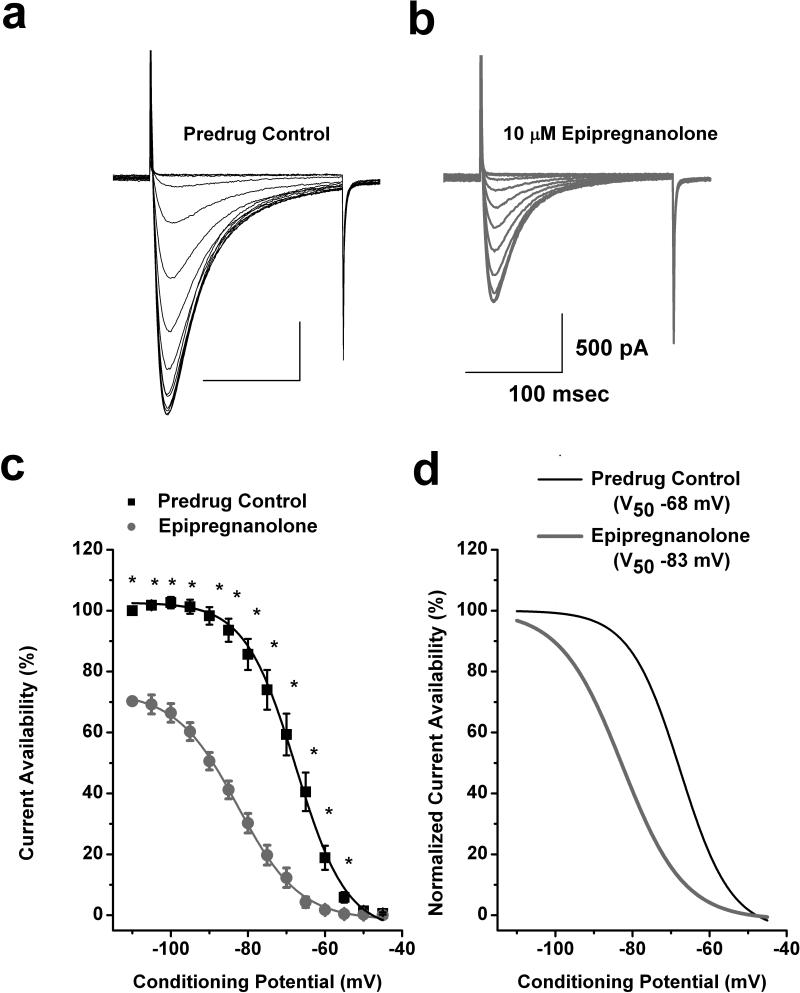
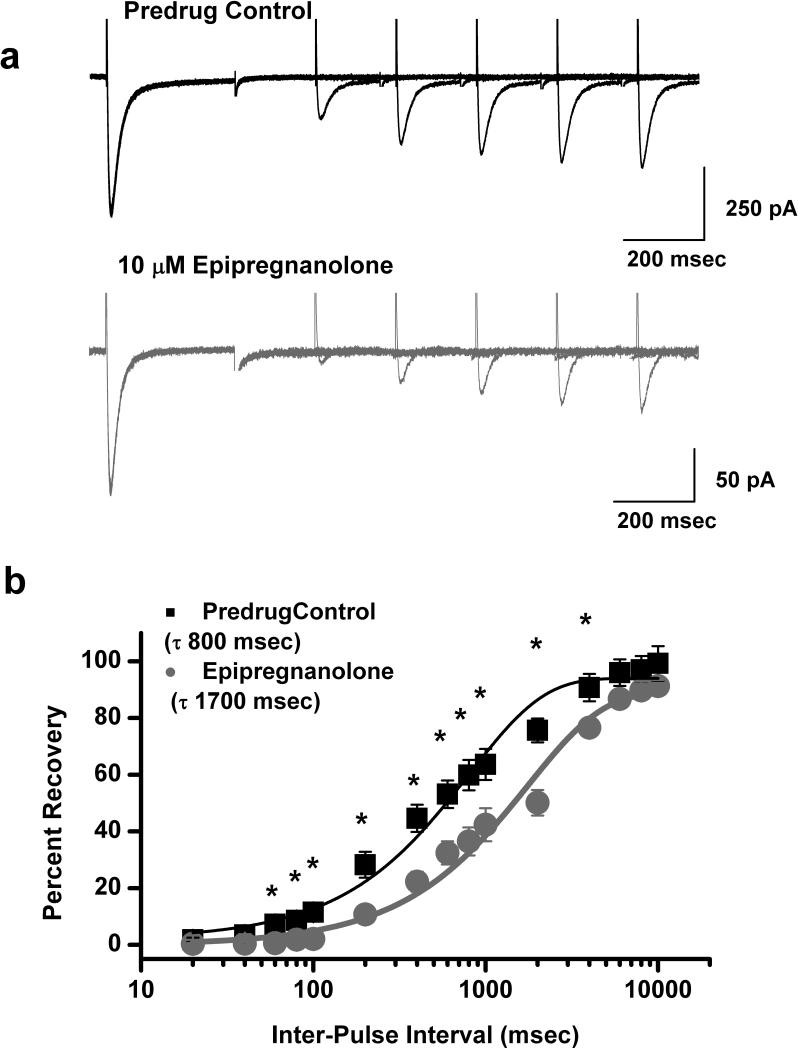
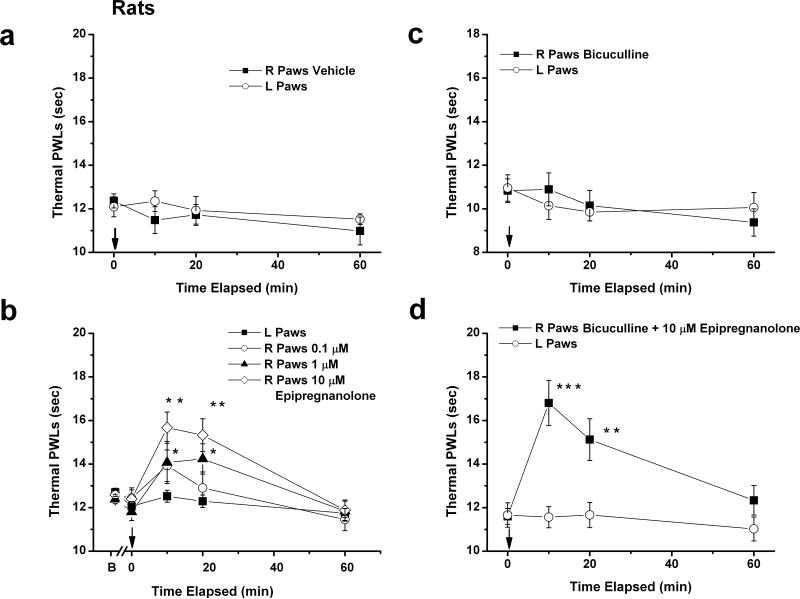
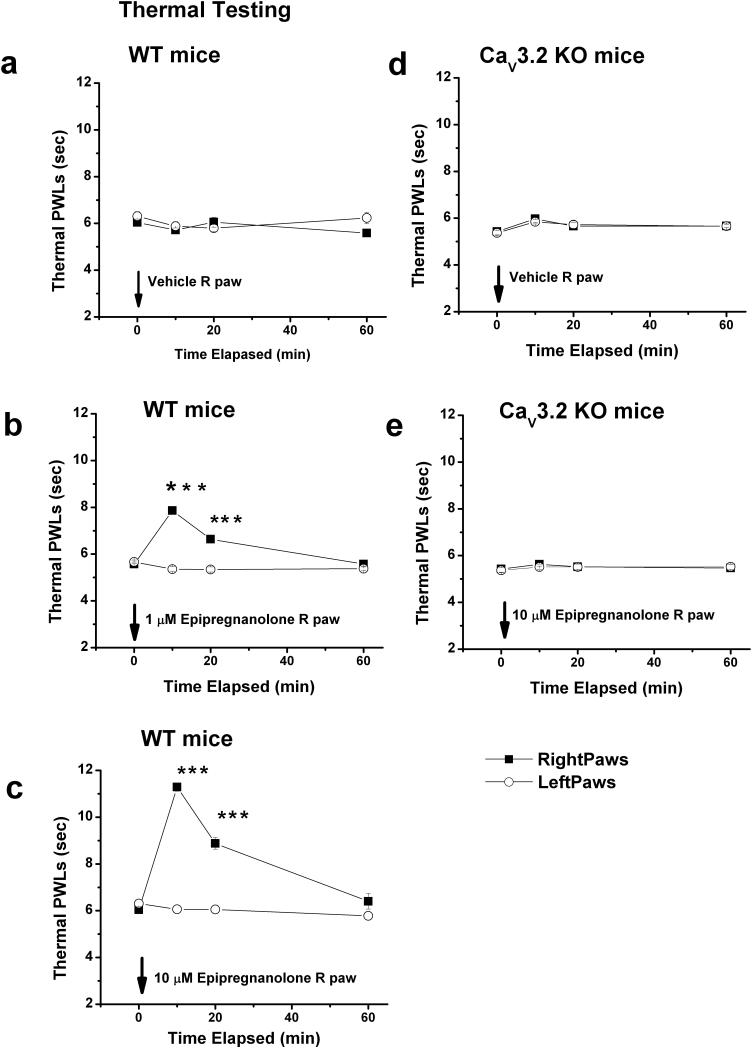
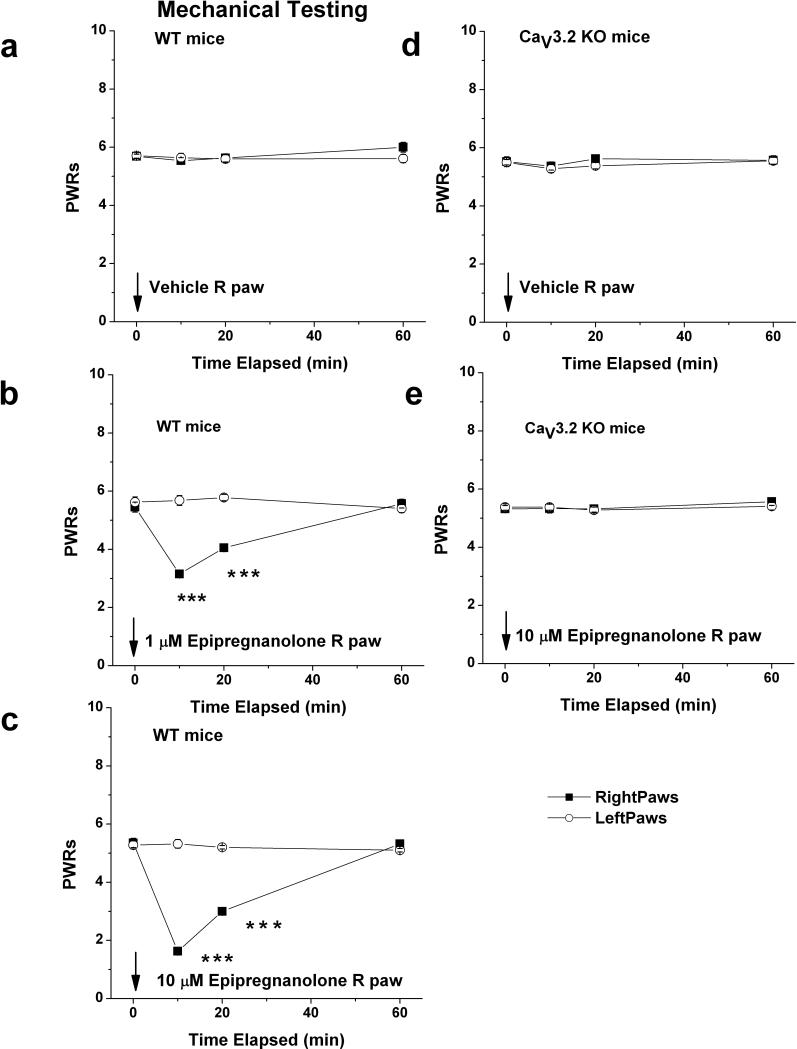
Results: We found that epipregnanolone reversibly blocked DRG T-currents with a half-maximal inhibitory concentration (IC50) of 2 μM and stabilized the channel in the inactive state. However, sodium, potassium, and gamma-aminobutyric acid (GABA)-gated ionic currents were not sensitive to the blocking effects of epipregnanolone even at 10 μM. In ensuing in vivo studies, we found that intraplantar (i.pl.) injections of epipregnanolone directly into peripheral receptive fields reduced responses to nociceptive heat stimuli in rats in a dose-dependent fashion. Furthermore, i.pl. epipregnanolone injections effectively reduced responses to peripheral nociceptive thermal and mechanical stimuli in wild-type mice but had no effect on the responses of CaV3.2 knockout mice.
Conclusions: We conclude that the inhibition of peripheral CaV3.2 T-channels contributes to the potent analgesic effect of the endogenous steroid epipregnanolone.
Figures
References
-
- Bicikova M, Hill M, Ripova D, Mohr P, Hampl R. Determination of steroid metabolome as a possible tool for laboratory diagnosis of schizophrenia. J Steroid Biochem Mol Biol. 2013;133:77–83. - PubMed
-
- Chen CC, Lamping KG, Nuno DW, Barresi R, Prouty SJ, Lavoie JL, Cribbs LL, England SK, Sigmund CD, Weiss RM, Williamson RA, Hill JA, Campbell CP. Abnormal coronary function in mice deficient in alpha1H T-type Ca2+ channels. Science. 2003;302:1416–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
