

Gastrointestinal microbes interact with canine adipose-derived mesenchymal stem cells in vitro and enhance immunomodulatory functions
- PMID: 24803072
- PMCID: PMC4120524
- DOI: 10.1089/scd.2014.0128
Gastrointestinal microbes interact with canine adipose-derived mesenchymal stem cells in vitro and enhance immunomodulatory functions
Abstract
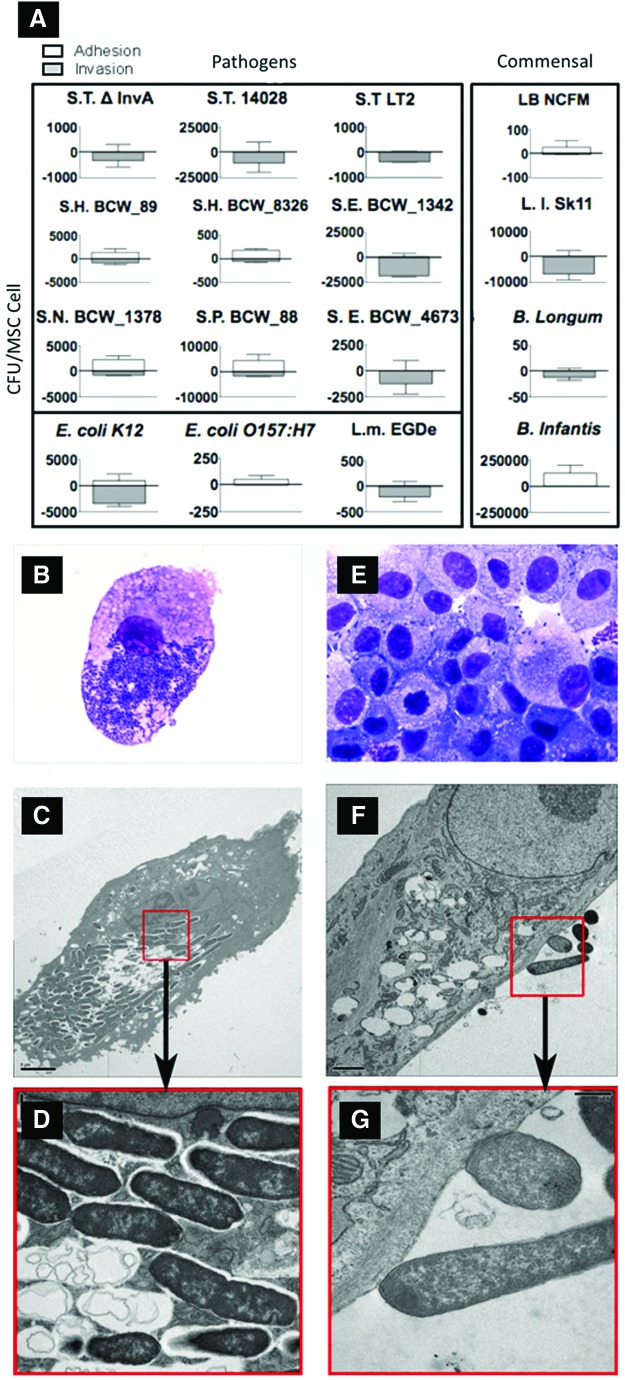
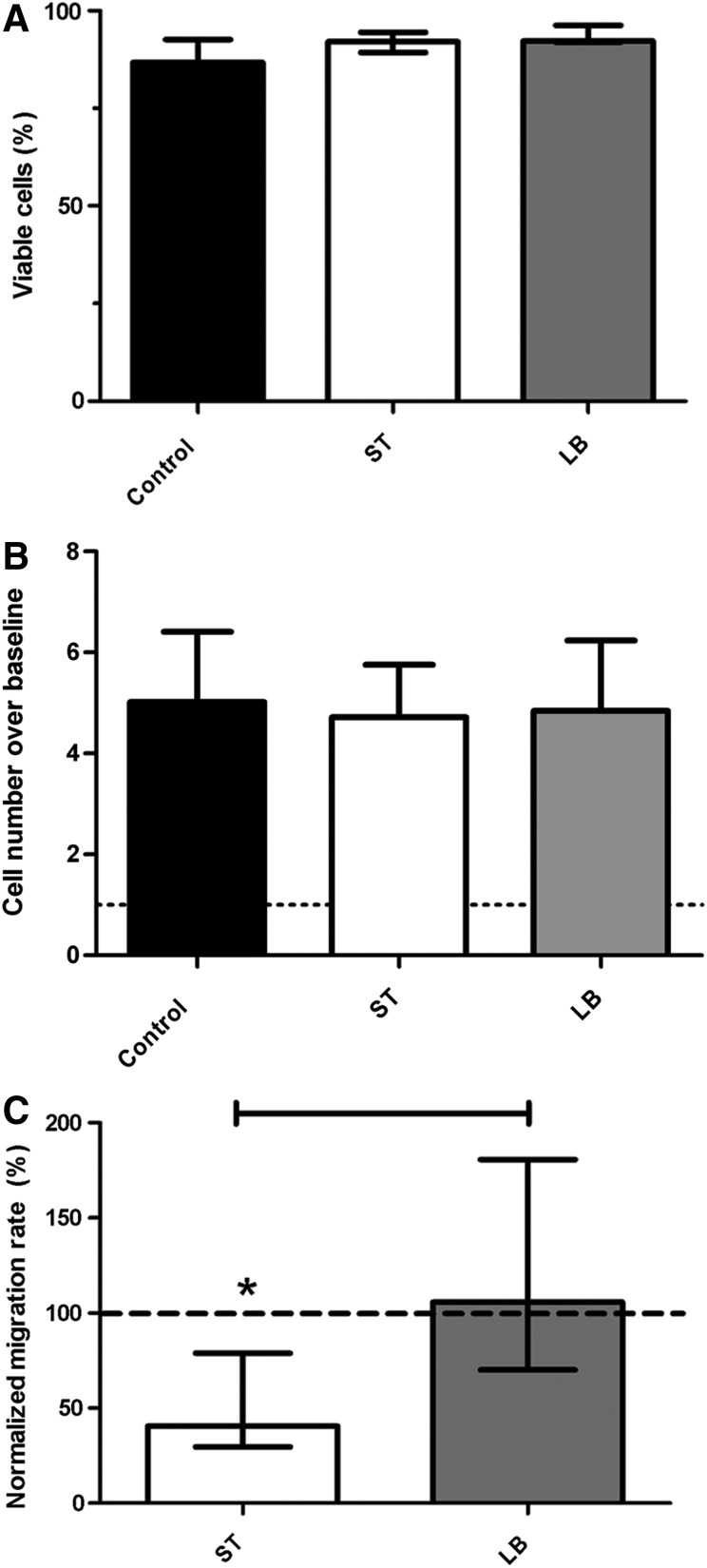
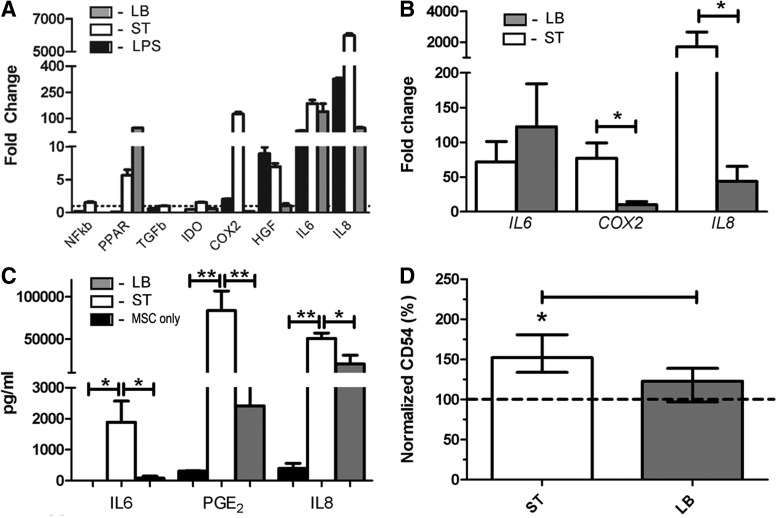
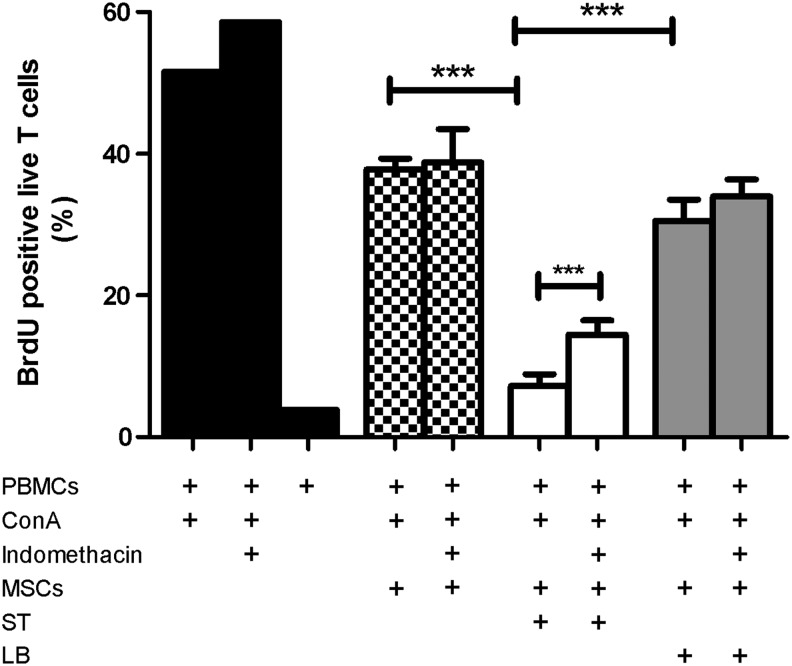
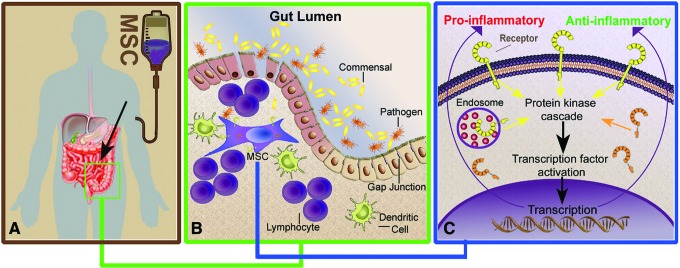
Mesenchymal stem cells (MSCs) are somatic, multipotent stromal cells with potent immunomodulatory and regenerative properties. Although MSCs have pattern recognition receptors and are modulated by Toll-like receptor ligands, MSC-microbial interactions are poorly defined. The objectives of this study were to determine the effect of bacterial association on MSC function. We hypothesized that gastrointestinal bacteria associate with MSCs and alter their immunomodulatory properties. The effect of MSC-microbial interactions on MSC morphology, viability, proliferation, migration, and immunomodulatory functions was investigated. MSCs associated with a remarkable array of enteric pathogens and commensal bacteria. MSC interactions with two model organisms, the pathogen Salmonella typhimurium and the probiotic Lactobacillus acidophilus, were further investigated. While ST readily invaded MSCs, LB adhered to the MSC plasma membrane. Neither microbe induced MSC death, degeneration, or diminished proliferation. Microbial association did not upregulate MHC-II, CD80/86, or CD1 expression. MSC-microbial interaction significantly increased transcription of key immunomodulatory genes, including COX2, IL6, and IL8, coupled with significantly increased prostaglandin E2 (PGE2), interleukin (IL)6, and IL8 secretion. MSC-ST coincubation resulted in increased MSC expression of CD54, and significant augmentation of MSC inhibition of mitogen-induced T-cell proliferation. T-cell proliferation was partially restored when PGE2 secretion was blocked from ST-primed MSCs. MSC-microbe interactions have a profound effect on MSC function and may be pivotal in a variety of clinical settings where MSCs are being explored as potential therapeutics in the context of microbial communities, such as Crohn's disease, chronic nonhealing wounds, and sepsis.
Figures
Similar articles
-
Influence of Donor's Age on Immunomodulatory Properties of Canine Adipose Tissue-Derived Mesenchymal Stem Cells.Stem Cells Dev. 2019 Dec 1;28(23):1562-1571. doi: 10.1089/scd.2019.0118. Epub 2019 Nov 11. Stem Cells Dev. 2019. PMID: 31588862
-
Comparative immunomodulatory properties of mesenchymal stem cells derived from human breast tumor and normal breast adipose tissue.Cancer Immunol Immunother. 2020 Sep;69(9):1841-1854. doi: 10.1007/s00262-020-02567-y. Epub 2020 Apr 30. Cancer Immunol Immunother. 2020. PMID: 32350594 Free PMC article.
-
Canine and Equine Mesenchymal Stem Cells Grown in Serum Free Media Have Altered Immunophenotype.Stem Cell Rev Rep. 2016 Apr;12(2):245-56. doi: 10.1007/s12015-015-9638-0. Stem Cell Rev Rep. 2016. PMID: 26638159 Free PMC article.
-
Immunosuppressive properties of mesenchymal stem cells: advances and applications.Curr Mol Med. 2012 Jun;12(5):574-91. doi: 10.2174/156652412800619950. Curr Mol Med. 2012. PMID: 22515979 Review.
-
Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential.Trends Pharmacol Sci. 2020 Sep;41(9):653-664. doi: 10.1016/j.tips.2020.06.009. Epub 2020 Jul 22. Trends Pharmacol Sci. 2020. PMID: 32709406 Free PMC article. Review.
Cited by
-
Therapeutic potential of mesenchymal stem cells for fungal infections: mechanisms, applications, and challenges.Front Microbiol. 2025 Jan 30;16:1554917. doi: 10.3389/fmicb.2025.1554917. eCollection 2025. Front Microbiol. 2025. PMID: 39949625 Free PMC article. Review.
-
Mechanisms utilized by feline adipose-derived mesenchymal stem cells to inhibit T lymphocyte proliferation.Stem Cell Res Ther. 2019 Jun 25;10(1):188. doi: 10.1186/s13287-019-1300-3. Stem Cell Res Ther. 2019. PMID: 31238978 Free PMC article.
-
Characterization and Immunomodulatory Effects of Canine Adipose Tissue- and Bone Marrow-Derived Mesenchymal Stromal Cells.PLoS One. 2016 Dec 1;11(12):e0167442. doi: 10.1371/journal.pone.0167442. eCollection 2016. PLoS One. 2016. PMID: 27907211 Free PMC article.
-
Immunosuppressive effect of mesenchymal stem cells on lung and gut CD8+ T cells in lipopolysaccharide-induced acute lung injury in mice.Cell Prolif. 2021 May;54(5):e13028. doi: 10.1111/cpr.13028. Epub 2021 Mar 19. Cell Prolif. 2021. PMID: 33738881 Free PMC article.
-
ADSCs stimulated by VEGF-C alleviate intestinal inflammation via dual mechanisms of enhancing lymphatic drainage by a VEGF-C/VEGFR-3-dependent mechanism and inhibiting the NF-κB pathway by the secretome.Stem Cell Res Ther. 2022 Sep 5;13(1):448. doi: 10.1186/s13287-022-03132-3. Stem Cell Res Ther. 2022. PMID: 36064450 Free PMC article.
References
-
- Singer NG. and Caplan AI. (2011). Mesenchymal stem cells: mechanisms of inflammation. Annu Rev Pathol 6:457–478 - PubMed
-
- Soleymaninejadian E, Pramanik K. and Samadian E. (2012). Immunomodulatory properties of mesenchymal stem cells: cytokines and factors. Am J Reprod Immunol 67:1–8 - PubMed
-
- Ghannam S, Pene J, Torcy-Moquet G, Jorgensen C. and Yssel H. (2010). Mesenchymal stem cells inhibit human Th17 cell differentiation and function and induce a T regulatory cell phenotype. J Immunol 185:302–312 - PubMed
-
- National Institute of Health US. Mesenchymal stem cells clinical trials. Available at: http://clinicaltrials.gov/ct2/results?term=mesenchymal+stem+cell (Accessed 8th August2014)
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
