

Sertoli cells control peritubular myoid cell fate and support adult Leydig cell development in the prepubertal testis
- PMID: 24803659
- PMCID: PMC4011090
- DOI: 10.1242/dev.107029
Sertoli cells control peritubular myoid cell fate and support adult Leydig cell development in the prepubertal testis
Abstract
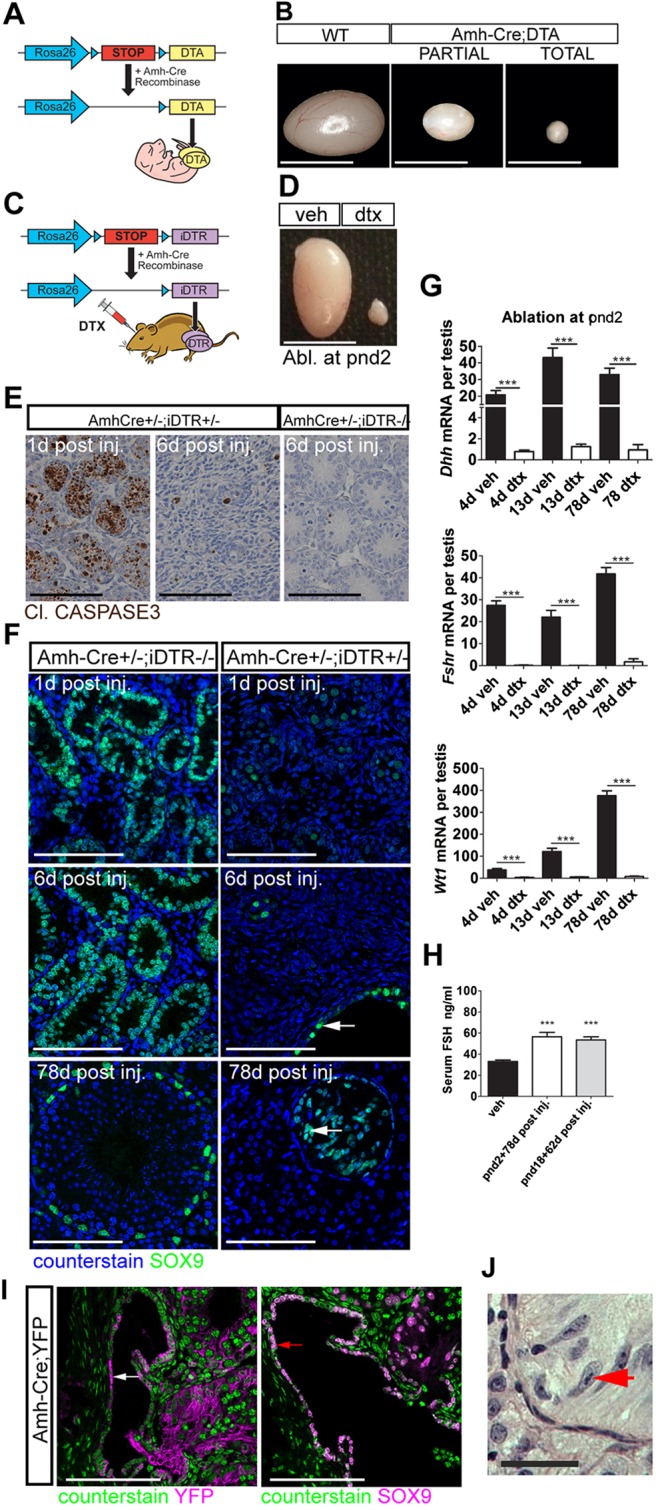
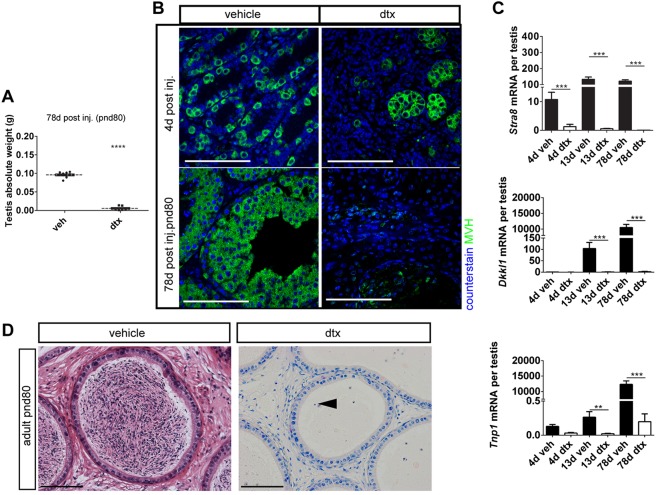
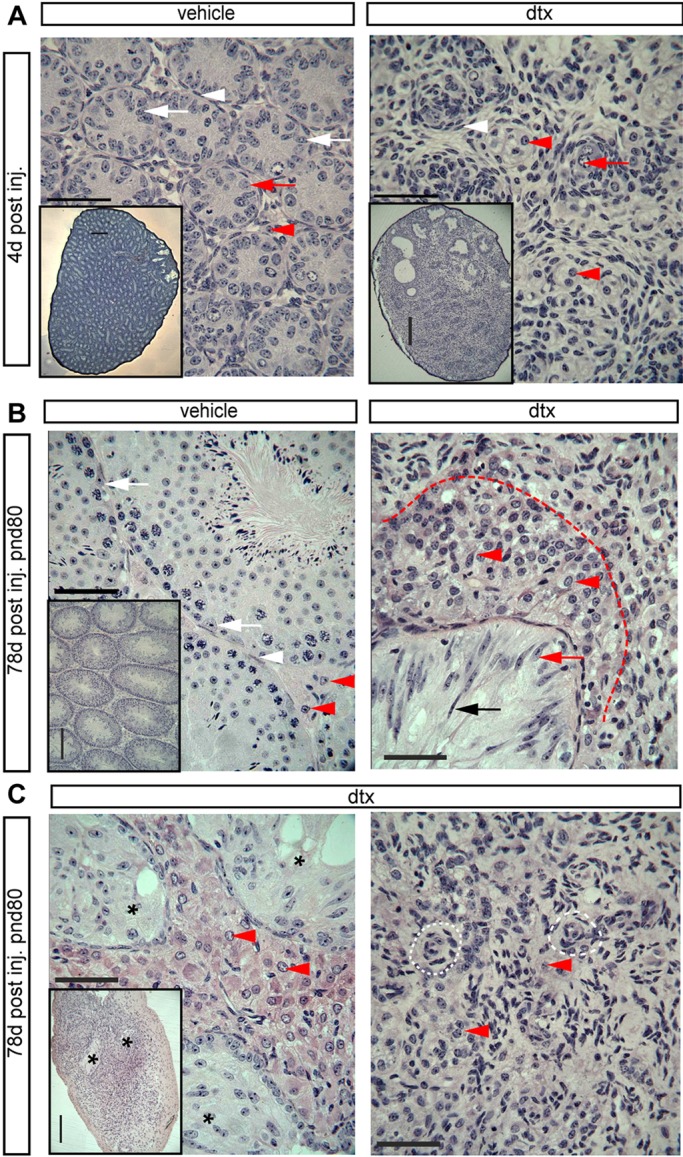
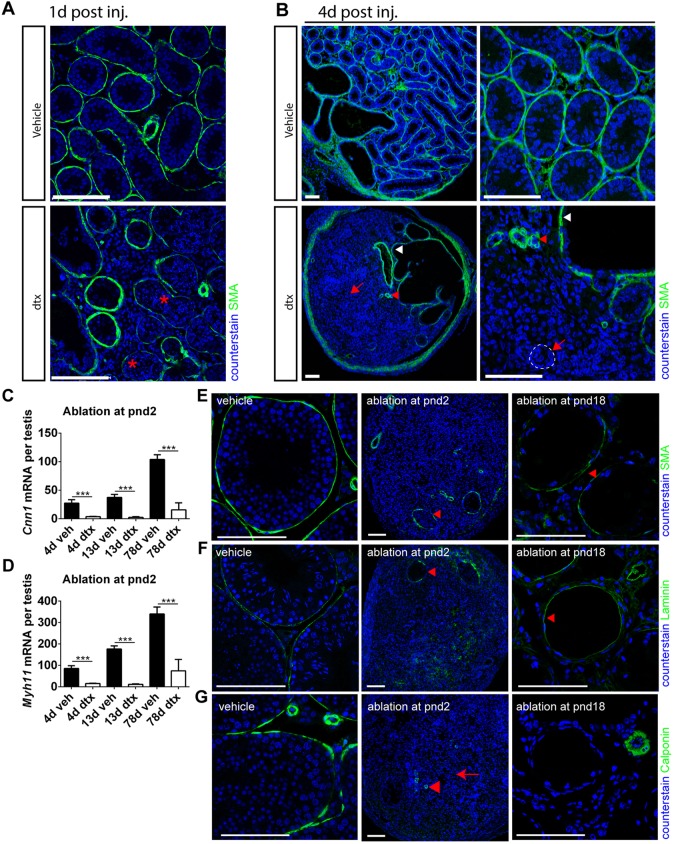
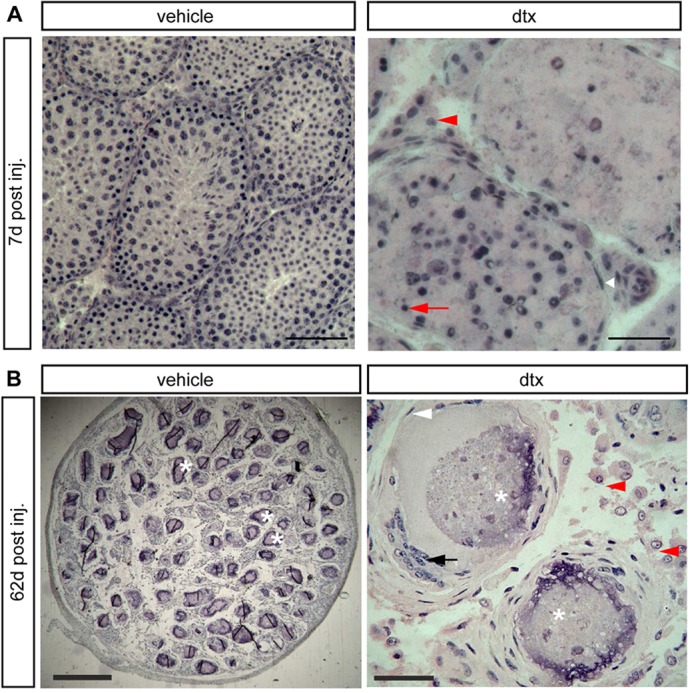
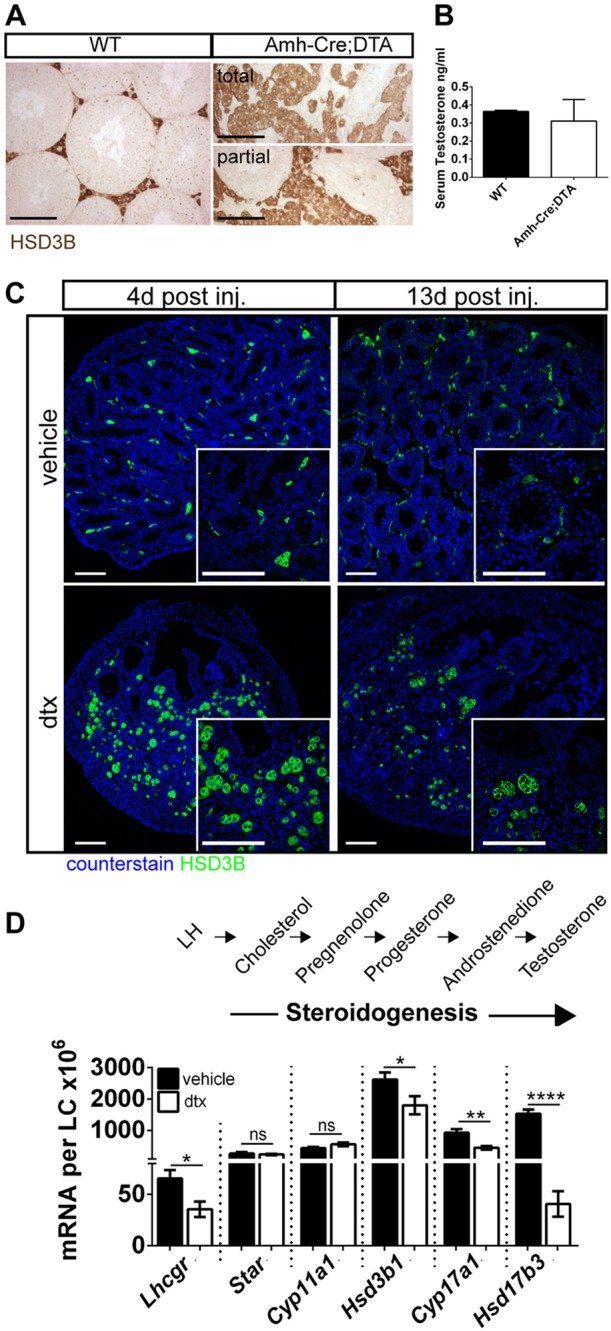
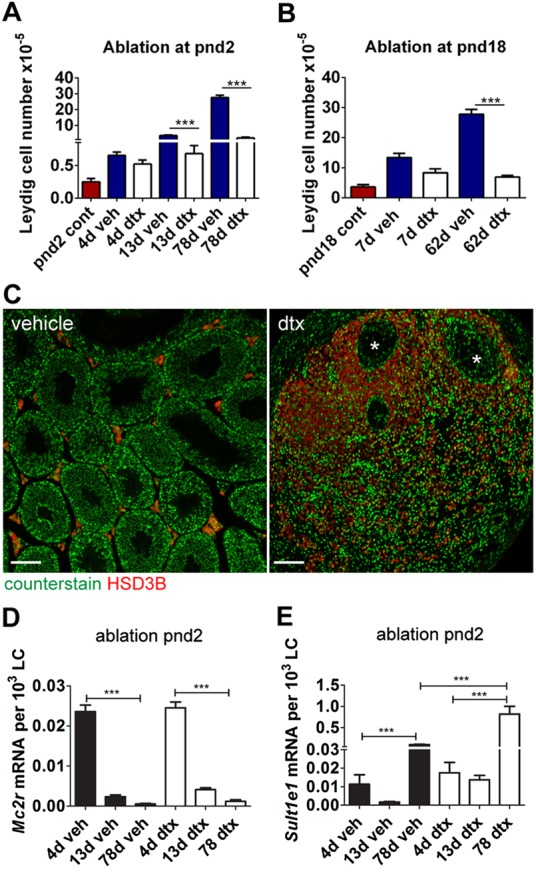
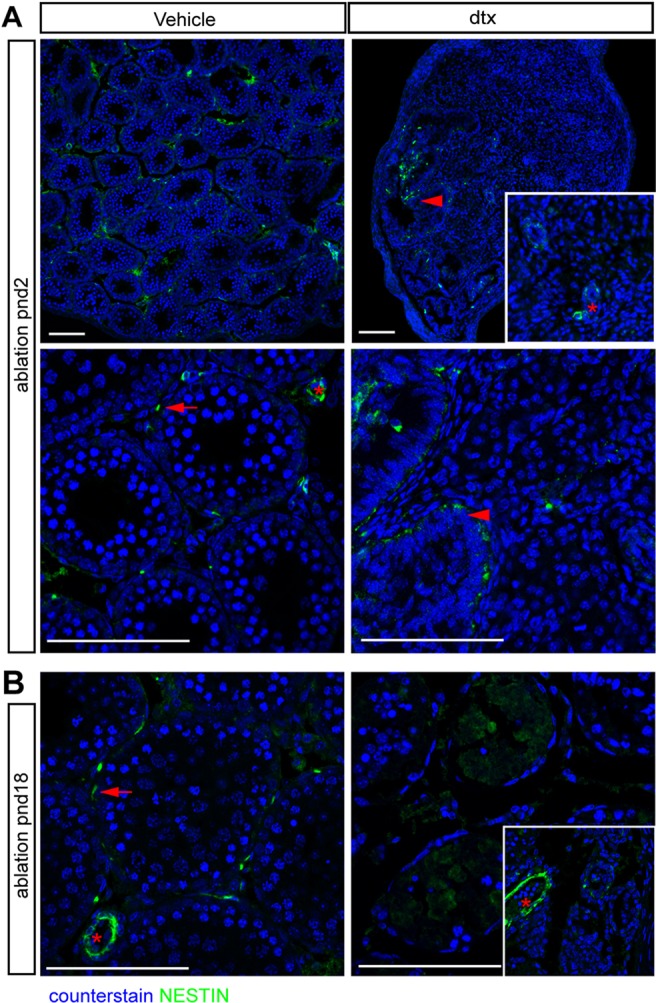
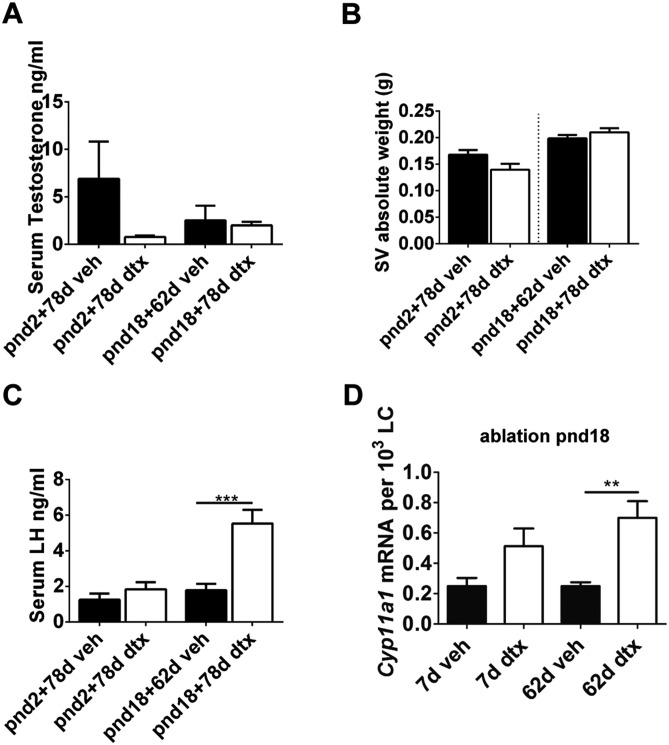
Sertoli cells (SCs) regulate testicular fate in the differentiating gonad and are the main regulators of spermatogenesis in the adult testis; however, their role during the intervening period of testis development, in particular during adult Leydig cell (ALC) differentiation and function, remains largely unknown. To examine SC function during fetal and prepubertal development we generated two transgenic mouse models that permit controlled, cell-specific ablation of SCs in pre- and postnatal life. Results show that SCs are required: (1) to maintain the differentiated phenotype of peritubular myoid cells (PTMCs) in prepubertal life; (2) to maintain the ALC progenitor population in the postnatal testis; and (3) for development of normal ALC numbers. Furthermore, our data show that fetal LCs function independently from SC, germ cell or PTMC support in the prepubertal testis. Together, these findings reveal that SCs remain essential regulators of testis development long after the period of sex determination. These findings have significant implications for our understanding of male reproductive disorders and wider androgen-related conditions affecting male health.
Keywords: Diphtheria; Leydig; Male fertility; Mouse; Peritubular myoid; Sertoli; Testis.
Figures
References
Publication types
MeSH terms
Grants and funding
- BB/JO1446X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/D/20211552/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G1002033/MRC_/Medical Research Council/United Kingdom
- G1100354/1/MRC_/Medical Research Council/United Kingdom
- G1100354/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
