

Glycosylinositol phosphorylceramides from Rosa cell cultures are boron-bridged in the plasma membrane and form complexes with rhamnogalacturonan II
- PMID: 24804932
- PMCID: PMC4230332
- DOI: 10.1111/tpj.12547
Glycosylinositol phosphorylceramides from Rosa cell cultures are boron-bridged in the plasma membrane and form complexes with rhamnogalacturonan II
Abstract
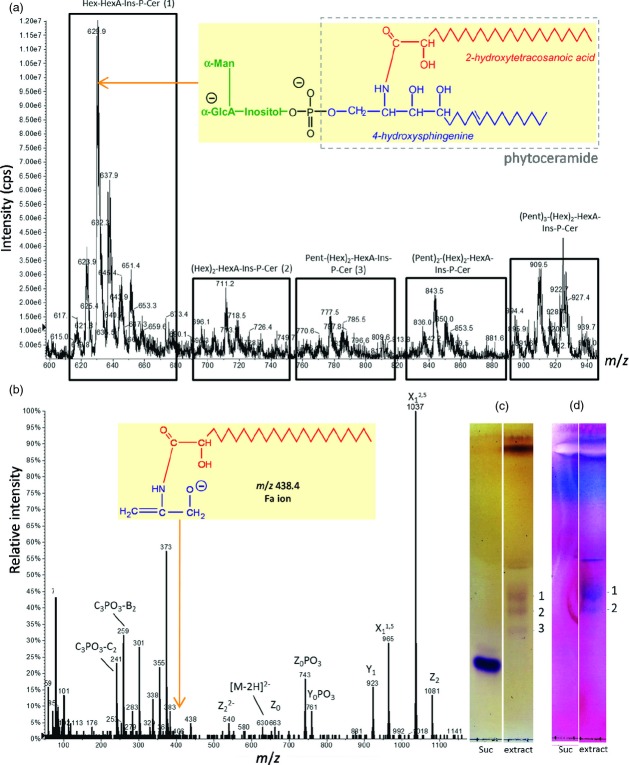
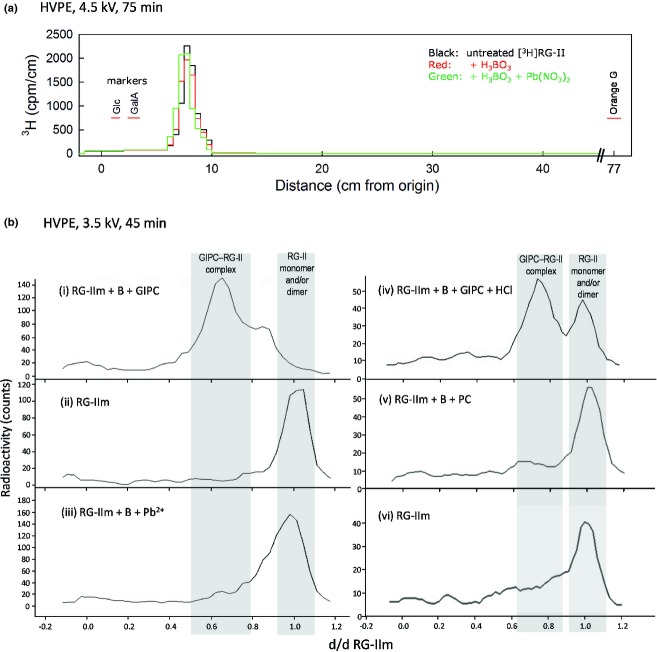
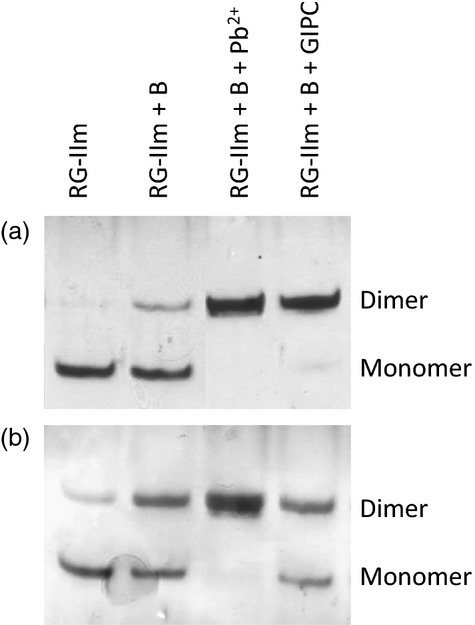
Boron (B) is essential for plant cell-wall structure and membrane functions. Compared with its role in cross-linking the pectic domain rhamnogalacturonan II (RG-II), little information is known about the biological role of B in membranes. Here, we investigated the involvement of glycosylinositol phosphorylceramides (GIPCs), major components of lipid rafts, in the membrane requirement for B. Using thin-layer chromatography and mass spectrometry, we first characterized GIPCs from Rosa cell culture. The major GIPC has one hexose residue, one hexuronic acid residue, inositol phosphate, and a ceramide moiety with a C18 trihydroxylated mono-unsaturated long-chain base and a C24 monohydroxylated saturated fatty acid. Disrupting B bridging (by B starvation in vivo or by treatment with cold dilute HCl or with excess borate in vitro) enhanced the GIPCs' extractability. As RG-II is the main B-binding site in plants, we investigated whether it could form a B-centred complex with GIPCs. Using high-voltage paper electrophoresis, we showed that addition of GIPCs decreased the electrophoretic mobility of radiolabelled RG-II, suggesting formation of a GIPC-B-RG-II complex. Last, using polyacrylamide gel electrophoresis, we showed that added GIPCs facilitate RG-II dimerization in vitro. We conclude that B plays a structural role in the plasma membrane. The disruption of membrane components by high borate may account for the phytotoxicity of excess B. Moreover, the in-vitro formation of a GIPC-B-RG-II complex gives the first molecular explanation of the wall-membrane attachment sites observed in vivo. Finally, our results suggest a role for GIPCs in the RG-II dimerization process.
Keywords: Rosa sp; boron; cell wall; cross-linking; glycosylinositol phosphoceramides; glycosylinositol phosphorylceramides; lipid rafts; plasma membrane; rhamnogalacturonan II.
© 2014 The Authors. The Plant Journal published by Society for Experimental Biology and John Wiley & Sons Ltd.
Figures


References
-
- Ahn J, Verma R, Kim M, Lee J, Kim Y, Bang J, Reiter W, Pai H. Depletion of UDP-d-apiose/UDP-d-xylose synthases results in rhamnogalacturonan-II deficiency, cell-wall thickening, and cell death in higher plants. J. Biol. Chem. 2006;281:13708–13716. - PubMed
-
- Al-Mustafa WA, Falatah AM, El-Shall AA. Effect of excess boron fertilization on status and availability of boron in calcareous soils. Fertilizer Res. 1993;36:71–78.
-
- Aquea F, Federici F, Moscoso C, Vega A, Jullian P, Haseloff JIM, Arce-Johnson P. A molecular framework for the inhibition of Arabidopsis root growth in response to boron toxicity. Plant, Cell Environ. 2012;35:719–734. - PubMed
-
- Blevins DG, Lukaszewski KM. Boron in plant structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998;49:481–500. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
