

The intriguing plant nuclear lamina
- PMID: 24808902
- PMCID: PMC4010787
- DOI: 10.3389/fpls.2014.00166
The intriguing plant nuclear lamina
Abstract
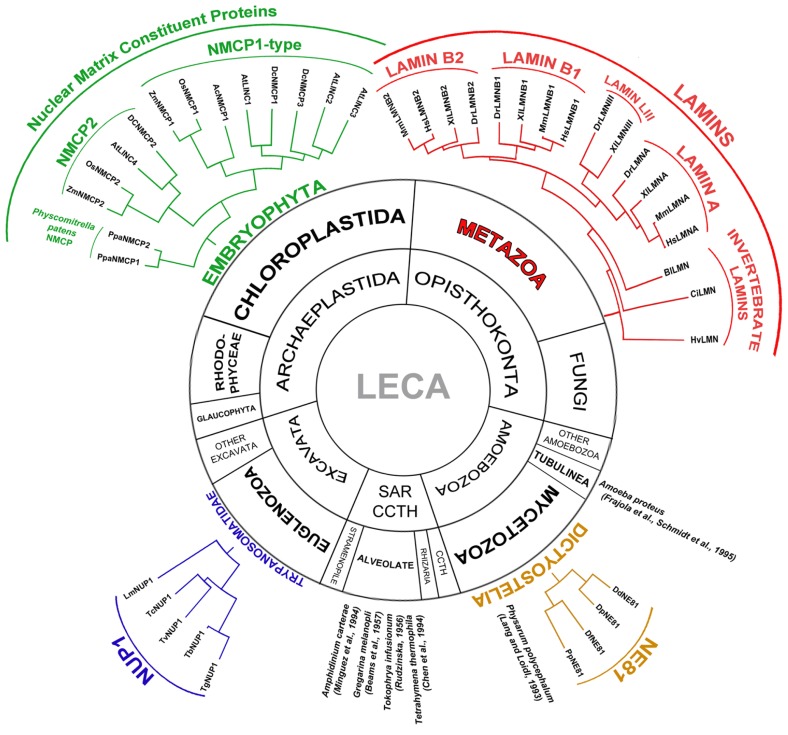
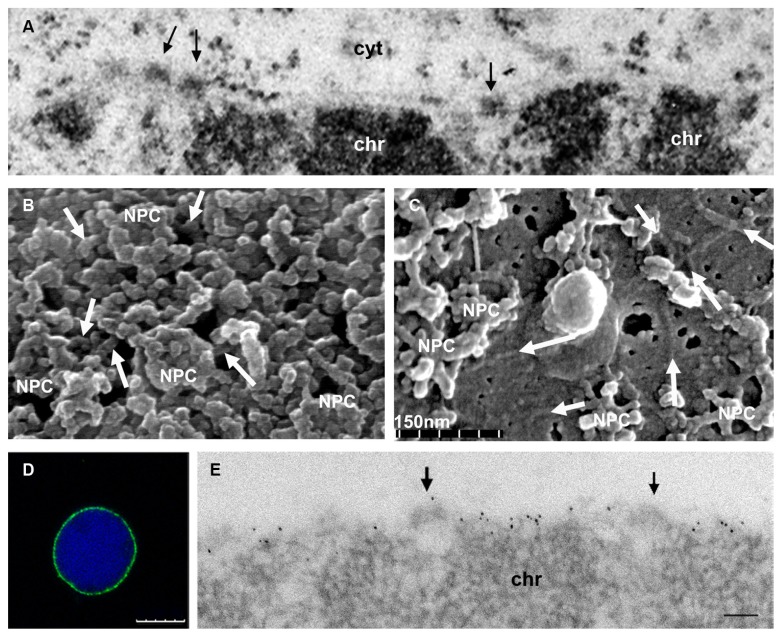
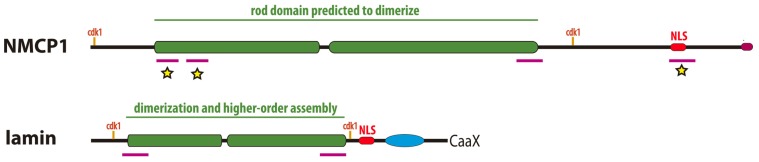
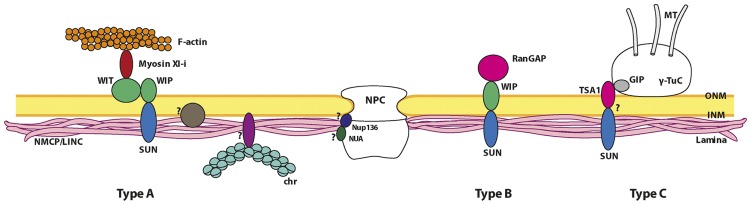
The nuclear lamina is a complex protein mesh attached to the inner nuclear membrane (INM), which is also associated with nuclear pore complexes. It provides mechanical support to the nucleus and nuclear envelope, and as well as facilitating the connection of the nucleoskeleton to the cytoskeleton, it is also involved in chromatin organization, gene regulation, and signaling. In metazoans, the nuclear lamina consists of a polymeric layer of lamins and other interacting proteins responsible for its association with the INM and chromatin. In plants, field emission scanning electron microscopy of nuclei, and thin section transmission electron microscopy of isolated nucleoskeletons, reveals the lamina to have a similar structure to that of metazoans. Moreover, although plants lack lamin genes and the genes encoding most lamin-binding proteins, the main functions of the lamina are fulfilled in plants. Hence, it would appear that the plant lamina is not based on lamins and that other proteins substitute for lamins in plant cells. The nuclear matrix constituent proteins are the best characterized structural proteins in the plant lamina. Although these proteins do not display strong sequence similarity to lamins, their predicted secondary structure and sub-nuclear distribution, as well as their influence on nuclear size and shape, and on heterochromatin organization, suggest they could be functional lamin analogs. In this review we shall summarize what is currently known about the organization and composition of the plant nuclear lamina and its interacting complexes, and we will discuss the activity of this structure in the plant cell and its nucleus.
Keywords: CRWN proteins; LINC proteins; NMCP proteins; Nup136; SUN proteins; plant nuclear envelope; plant nuclear lamina; plant nucleocytoplasmic linkers.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources