

A dominant-negative mutation of mouse Lmx1b causes glaucoma and is semi-lethal via LDB1-mediated dimerization [corrected]
- PMID: 24809698
- PMCID: PMC4014447
- DOI: 10.1371/journal.pgen.1004359
A dominant-negative mutation of mouse Lmx1b causes glaucoma and is semi-lethal via LDB1-mediated dimerization [corrected]
Erratum in
- PLoS Genet. 2014 Dec;10(12):e1004917
Abstract
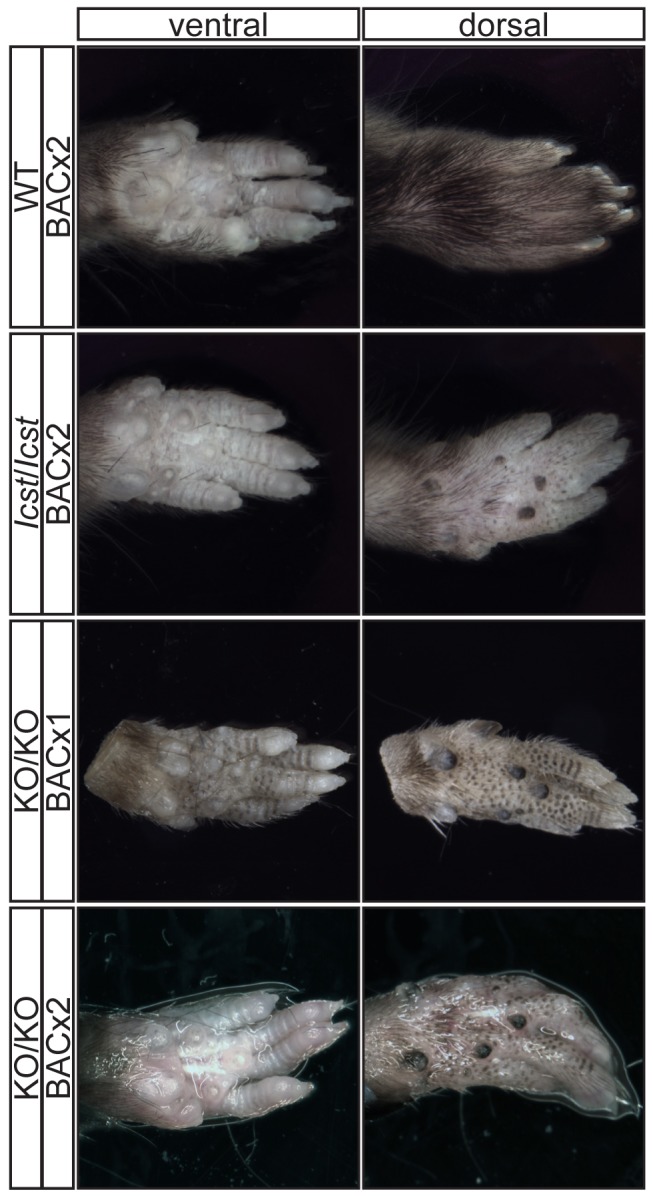
Mutations in the LIM-homeodomain transcription factor LMX1B cause nail-patella syndrome, an autosomal dominant pleiotrophic human disorder in which nail, patella and elbow dysplasia is associated with other skeletal abnormalities and variably nephropathy and glaucoma. It is thought to be a haploinsufficient disorder. Studies in the mouse have shown that during development Lmx1b controls limb dorsal-ventral patterning and is also required for kidney and eye development, midbrain-hindbrain boundary establishment and the specification of specific neuronal subtypes. Mice completely deficient for Lmx1b die at birth. In contrast to the situation in humans, heterozygous null mice do not have a mutant phenotype. Here we report a novel mouse mutant Icst, an N-ethyl-N-nitrosourea-induced missense substitution, V265D, in the homeodomain of LMX1B that abolishes DNA binding and thereby the ability to transactivate other genes. Although the homozygous phenotypic consequences of Icst and the null allele of Lmx1b are the same, heterozygous Icst elicits a phenotype whilst the null allele does not. Heterozygous Icst causes glaucomatous eye defects and is semi-lethal, probably due to kidney failure. We show that the null phenotype is rescued more effectively by an Lmx1b transgene than is Icst. Co-immunoprecipitation experiments show that both wild-type and Icst LMX1B are found in complexes with LIM domain binding protein 1 (LDB1), resulting in lower levels of functional LMX1B in Icst heterozygotes than null heterozygotes. We conclude that Icst is a dominant-negative allele of Lmx1b. These findings indicate a reassessment of whether nail-patella syndrome is always haploinsufficient. Furthermore, Icst is a rare example of a model of human glaucoma caused by mutation of the same gene in humans and mice.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Dreyer SD, Zhou G, Baldini A, Winterpacht A, Zabel B, et al. (1998) Mutations in LMX1B cause abnormal skeletal patterning and renal dysplasia in nail patella syndrome. Nat Genet 19: 47–50. - PubMed
-
- Vollrath D, Jaramillo-Babb VL, Clough MV, McIntosh I, Scott KM, et al. (1998) Loss-of-function mutations in the LIM-homeodomain gene, LMX1B, in nail-patella syndrome. Hum Mol Genet 7: 1091–1098. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
