

Insights into the architecture of the eIF2Bα/β/δ regulatory subcomplex
- PMID: 24811713
- PMCID: PMC4045321
- DOI: 10.1021/bi500346u
Insights into the architecture of the eIF2Bα/β/δ regulatory subcomplex
Abstract
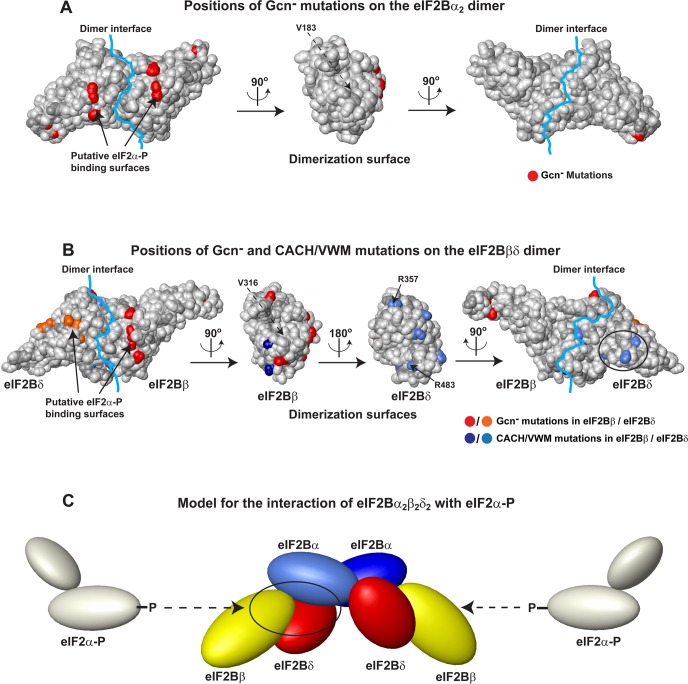
Eukaryotic translation initiation factor 2B (eIF2B), the guanine nucleotide exchange factor for the G-protein eIF2, is one of the main targets for the regulation of protein synthesis. The eIF2B activity is inhibited in response to a wide range of stress factors and diseases, including viral infections, hypoxia, nutrient starvation, and heme deficiency, collectively known as the integrated stress response. eIF2B has five subunits (α-ε). The α, β, and δ subunits are homologous to each other and form the eIF2B regulatory subcomplex, which is believed to be a trimer consisting of monomeric α, β, and δ subunits. Here we use a combination of biophysical methods, site-directed mutagenesis, and bioinformatics to show that the human eIF2Bα subunit is in fact a homodimer, at odds with the current trimeric model for the eIF2Bα/β/δ regulatory complex. eIF2Bα dimerizes using the same interface that is found in the homodimeric archaeal eIF2Bα/β/δ homolog aIF2B and related metabolic enzymes. We also present evidence that the eIF2Bβ/δ binding interface is similar to that in the eIF2Bα2 homodimer. Mutations at the predicted eIF2Bβ/δ dimer interface cause genetic neurological disorders in humans. We propose that the eIF2B regulatory subcomplex is an α2β2δ2 hexamer, composed of one α2 homodimer and two βδ heterodimers. Our results offer novel insights into the architecture of eIF2B and its interactions with the G-protein eIF2.
Figures





Similar articles
-
Architecture of the eIF2B regulatory subcomplex and its implications for the regulation of guanine nucleotide exchange on eIF2.Nucleic Acids Res. 2015 Nov 16;43(20):9994-10014. doi: 10.1093/nar/gkv930. Epub 2015 Sep 17. Nucleic Acids Res. 2015. PMID: 26384431 Free PMC article.
-
Archaeal aIF2B interacts with eukaryotic translation initiation factors eIF2alpha and eIF2Balpha: Implications for aIF2B function and eIF2B regulation.J Mol Biol. 2009 Sep 25;392(3):701-22. doi: 10.1016/j.jmb.2009.07.030. Epub 2009 Jul 17. J Mol Biol. 2009. PMID: 19616556 Free PMC article.
-
Crystal structure of the alpha subunit of human translation initiation factor 2B.J Mol Biol. 2009 Oct 2;392(4):937-51. doi: 10.1016/j.jmb.2009.07.054. Epub 2009 Jul 23. J Mol Biol. 2009. PMID: 19631657
-
eIF2B: recent structural and functional insights into a key regulator of translation.Biochem Soc Trans. 2015 Dec;43(6):1234-40. doi: 10.1042/BST20150164. Biochem Soc Trans. 2015. PMID: 26614666 Review.
-
Crystal structure of eIF2B and insights into eIF2-eIF2B interactions.FEBS J. 2017 Mar;284(6):868-874. doi: 10.1111/febs.13896. Epub 2016 Sep 29. FEBS J. 2017. PMID: 27627185 Review.
Cited by
-
eIF2Bδ blocks the integrated stress response and maintains eIF2B activity and cancer metastasis by overexpression in breast cancer stem cells.Proc Natl Acad Sci U S A. 2023 Apr 11;120(15):e2207898120. doi: 10.1073/pnas.2207898120. Epub 2023 Apr 4. Proc Natl Acad Sci U S A. 2023. PMID: 37014850 Free PMC article.
-
Regulation and function of elF2B in neurological and metabolic disorders.Biosci Rep. 2022 Jun 30;42(6):BSR20211699. doi: 10.1042/BSR20211699. Biosci Rep. 2022. PMID: 35579296 Free PMC article. Review.
-
Molecular glues for protein-protein interactions: Progressing toward a new dream.Cell Chem Biol. 2024 Jun 20;31(6):1064-1088. doi: 10.1016/j.chembiol.2024.04.002. Epub 2024 May 2. Cell Chem Biol. 2024. PMID: 38701786 Free PMC article. Review.
-
Stress responses. Mutations in a translation initiation factor identify the target of a memory-enhancing compound.Science. 2015 May 29;348(6238):1027-30. doi: 10.1126/science.aaa6986. Epub 2015 Apr 9. Science. 2015. PMID: 25858979 Free PMC article.
-
Characterization and Mapping of retr04, retr05 and retr06 Broad-Spectrum Resistances to Turnip Mosaic Virus in Brassica juncea, and the Development of Robust Methods for Utilizing Recalcitrant Genotyping Data.Front Plant Sci. 2022 Jan 12;12:787354. doi: 10.3389/fpls.2021.787354. eCollection 2021. Front Plant Sci. 2022. PMID: 35095961 Free PMC article.
References
-
- Marintchev A.; Wagner G. (2004) Translation initiation: Structures, mechanisms and evolution. Q. Rev. Biophys. 37, 197–284. - PubMed
-
- Pestova T. V., Lorsch J. R., and Hellen C. U. T. (2007) The Mechanism of Translation Initiation in Eukaryotes. In Translational Control in Biology and Medicine (Mathews M. B., Sonenberg N., and Hershey J. W. B., Eds.) pp 87–128, Cold Spring Harbor Laboratory Press, Plainview, NY.
-
- Moreno J. A.; Halliday M.; Molloy C.; Radford H.; Verity N.; Axten J. M.; Ortori C. A.; Willis A. E.; Fischer P. M.; Barrett D. A.; Mallucci G. R. (2013) Oral treatment targeting the unfolded protein response prevents neurodegeneration and clinical disease in prion-infected mice. Sci. Transl. Med. 5, 206ra138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
