

Spatially distributed local fields in the hippocampus encode rat position
- PMID: 24812401
- PMCID: PMC4909490
- DOI: 10.1126/science.1250444
Spatially distributed local fields in the hippocampus encode rat position
Abstract
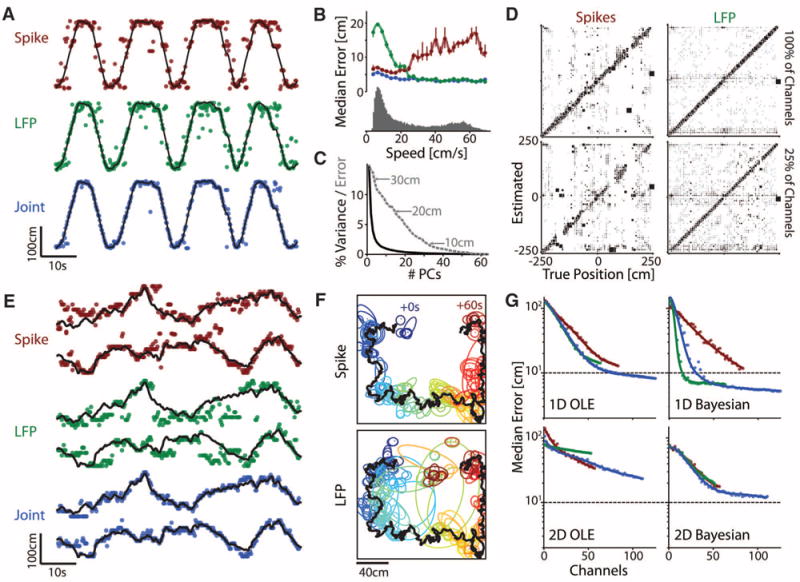
Although neuronal spikes can be readily detected from extracellular recordings, synaptic and subthreshold activity remains undifferentiated within the local field potential (LFP). In the hippocampus, neurons discharge selectively when the rat is at certain locations, while LFPs at single anatomical sites exhibit no such place-tuning. Nonetheless, because the representation of position is sparse and distributed, we hypothesized that spatial information can be recovered from multiple-site LFP recordings. Using high-density sampling of LFP and computational methods, we show that the spatiotemporal structure of the theta rhythm can encode position as robustly as neuronal spiking populations. Because our approach exploits the rhythmicity and sparse structure of neural activity, features found in many brain regions, it is useful as a general tool for discovering distributed LFP codes.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
