

Predictive polymer modeling reveals coupled fluctuations in chromosome conformation and transcription
- PMID: 24813616
- PMCID: PMC4427251
- DOI: 10.1016/j.cell.2014.03.025
Predictive polymer modeling reveals coupled fluctuations in chromosome conformation and transcription
Abstract
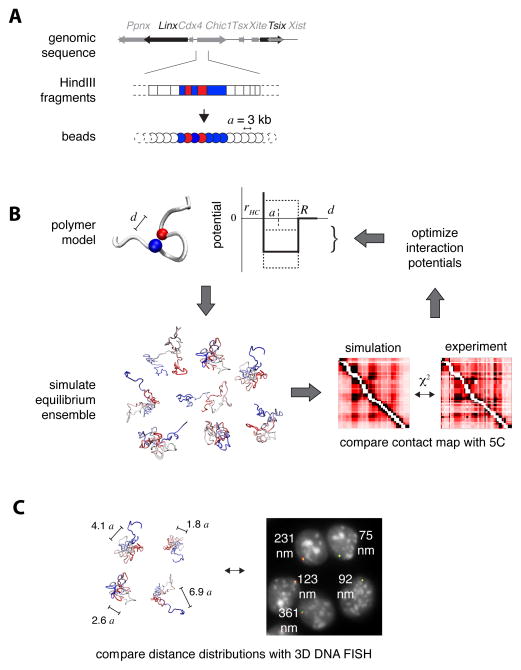
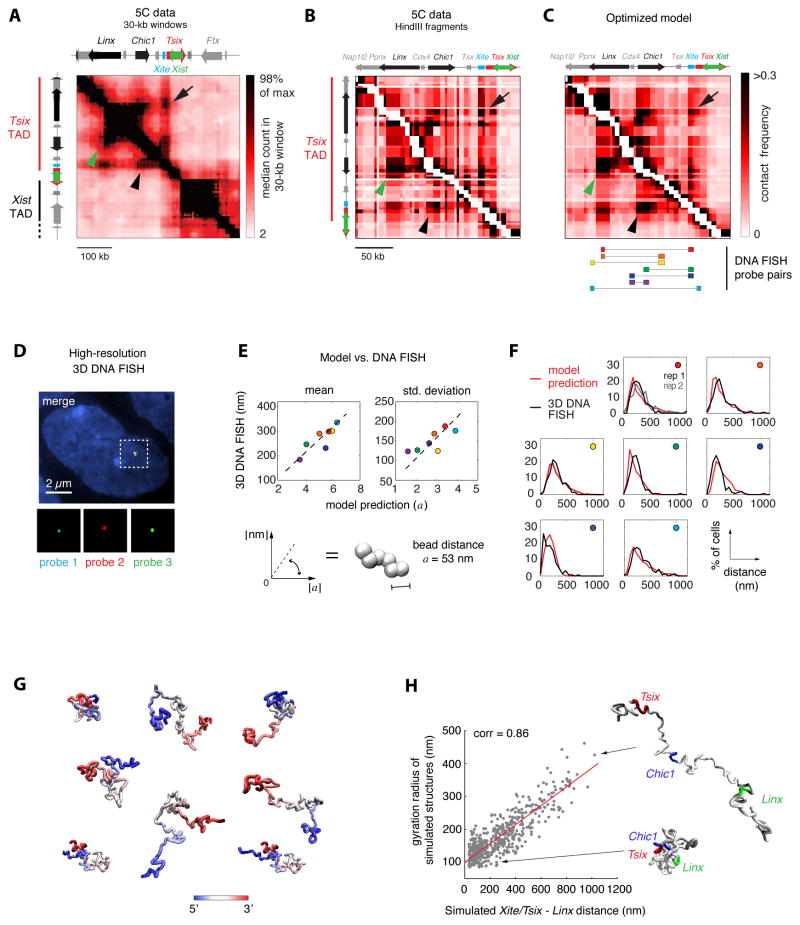
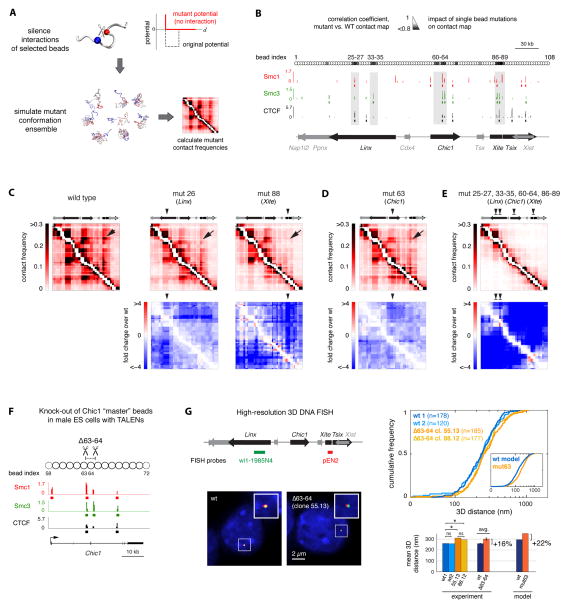
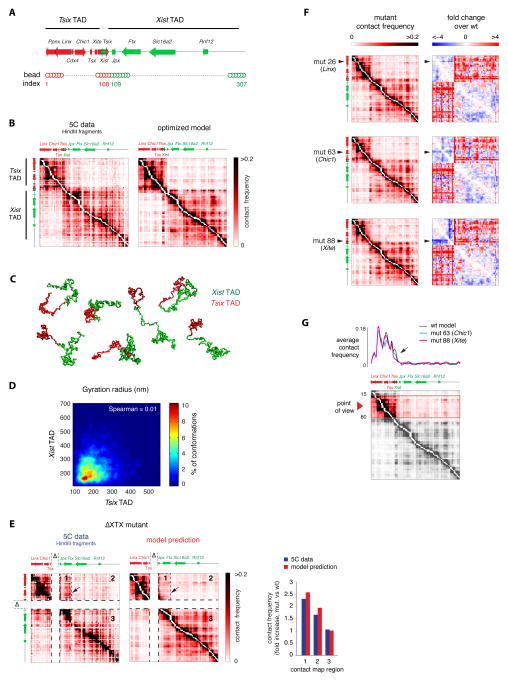
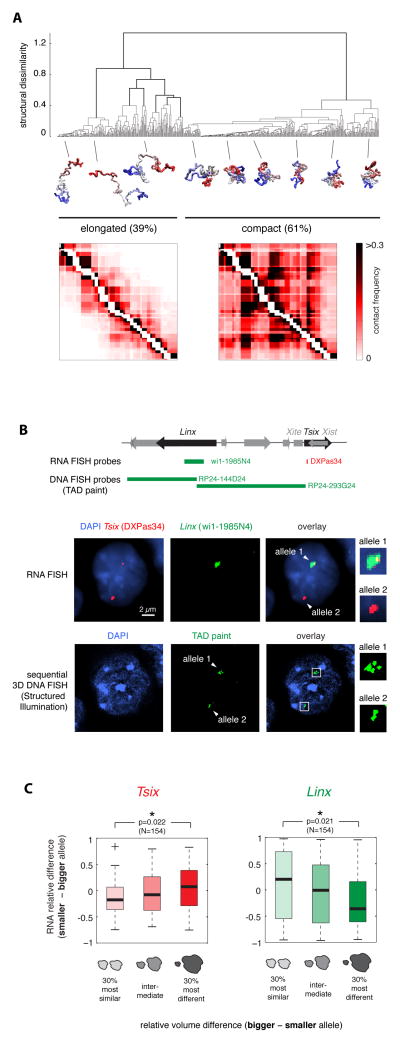
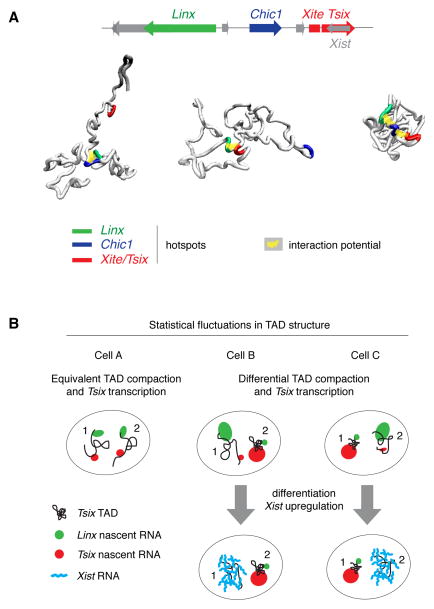
A new level of chromosome organization, topologically associating domains (TADs), was recently uncovered by chromosome conformation capture (3C) techniques. To explore TAD structure and function, we developed a polymer model that can extract the full repertoire of chromatin conformations within TADs from population-based 3C data. This model predicts actual physical distances and to what extent chromosomal contacts vary between cells. It also identifies interactions within single TADs that stabilize boundaries between TADs and allows us to identify and genetically validate key structural elements within TADs. Combining the model's predictions with high-resolution DNA FISH and quantitative RNA FISH for TADs within the X-inactivation center (Xic), we dissect the relationship between transcription and spatial proximity to cis-regulatory elements. We demonstrate that contacts between potential regulatory elements occur in the context of fluctuating structures rather than stable loops and propose that such fluctuations may contribute to asymmetric expression in the Xic during X inactivation.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Chromosomes: Dynamically in the loop.Nat Rev Genet. 2014 Jul;15(7):440. doi: 10.1038/nrg3762. Epub 2014 May 20. Nat Rev Genet. 2014. PMID: 24840553 No abstract available.
References
-
- Amano T, Sagai T, Tanabe H, Mizushina Y, Nakazawa H, Shiroishi T. Chromosomal Dynamics at the Shh Locus: Limb Bud-Specific Differential Regulation of Competence and Active Transcription. Dev Cell. 2009;16:47–57. - PubMed
-
- Andrey G, Montavon T, Mascrez B, Gonzalez F, Noordermeer D, Leleu M, Trono D, Spitz F, Duboule D. A Switch Between Topological Domains Underlies HoxD Genes Collinearity in Mouse Limbs. Science. 2013;340:1234167. - PubMed
-
- Baù D, Marti-Renom MA. Structure determination of genomic domains by satisfaction of spatial restraints. Chromosome Res. 2010;19:25–35. - PubMed
-
- Chaumeil J, Augui S, Chow JC, Heard E. Combined Immunofluorescence, RNA Fluorescent In Situ Hybridization, and DNA Fluorescent In Situ Hybridization to Study Chromatin Changes, Transcriptional Activity, Nuclear Organization, and X-Chromosome Inactivation. In: Hancock R, editor. The Nucleus. Totowa, NJ: Humana Press; 2008. pp. 297–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
