

Isolation of chromatin from dysfunctional telomeres reveals an important role for Ring1b in NHEJ-mediated chromosome fusions
- PMID: 24813883
- PMCID: PMC4054697
- DOI: 10.1016/j.celrep.2014.04.002
Isolation of chromatin from dysfunctional telomeres reveals an important role for Ring1b in NHEJ-mediated chromosome fusions
Abstract
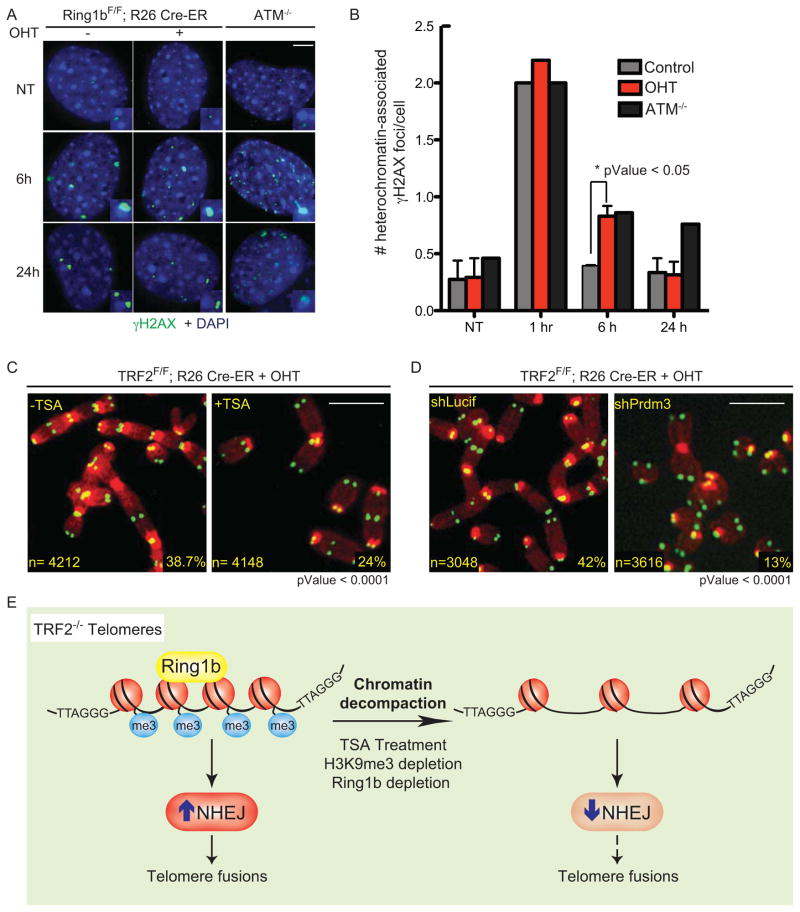
When telomeres become critically short, DNA damage response factors are recruited at chromosome ends, initiating a cellular response to DNA damage. We performed proteomic isolation of chromatin fragments (PICh) in order to define changes in chromatin composition that occur upon onset of acute telomere dysfunction triggered by depletion of the telomere-associated factor TRF2. This unbiased purification of telomere-associated proteins in functional or dysfunctional conditions revealed the dynamic changes in chromatin composition that take place at telomeres upon DNA damage induction. On the basis of our results, we describe a critical role for the polycomb group protein Ring1b in nonhomologous end-joining (NHEJ)-mediated end-to-end chromosome fusions. We show that cells with reduced levels of Ring1b have a reduced ability to repair uncapped telomeric chromatin. Our data represent an unbiased isolation of chromatin undergoing DNA damage and are a valuable resource to map the changes in chromatin composition in response to DNA damage activation.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
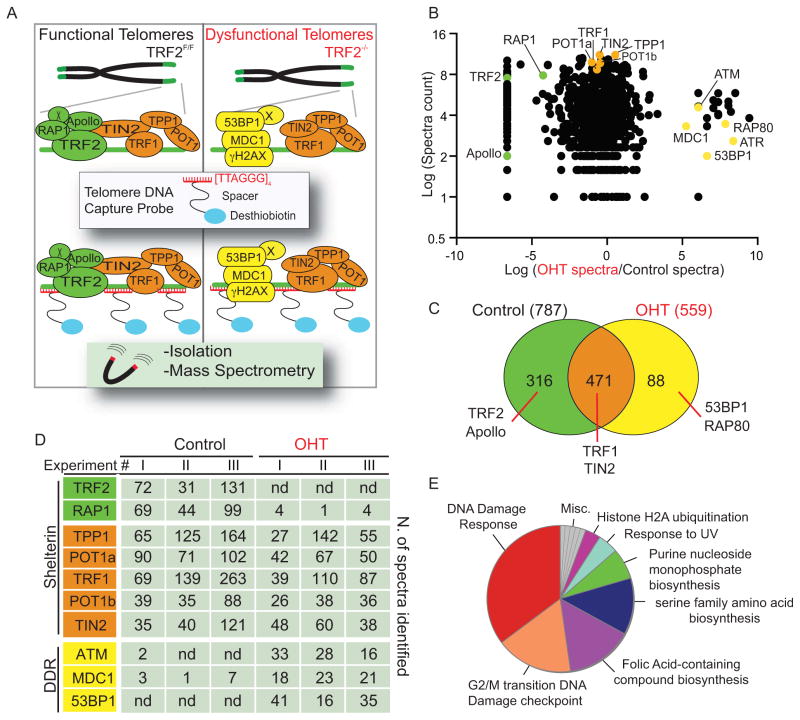
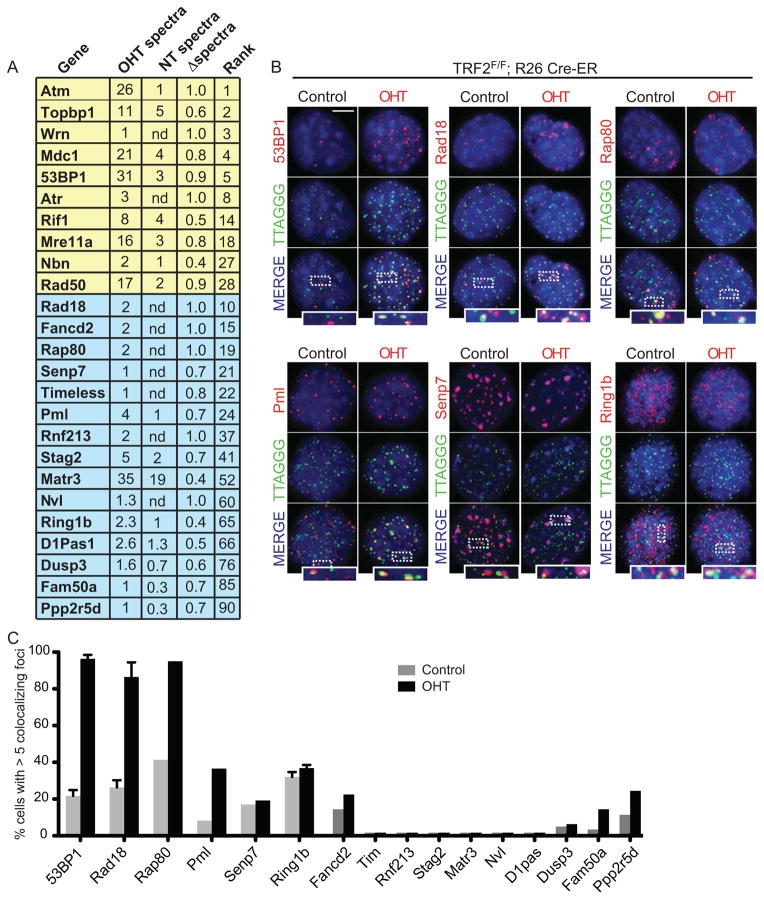
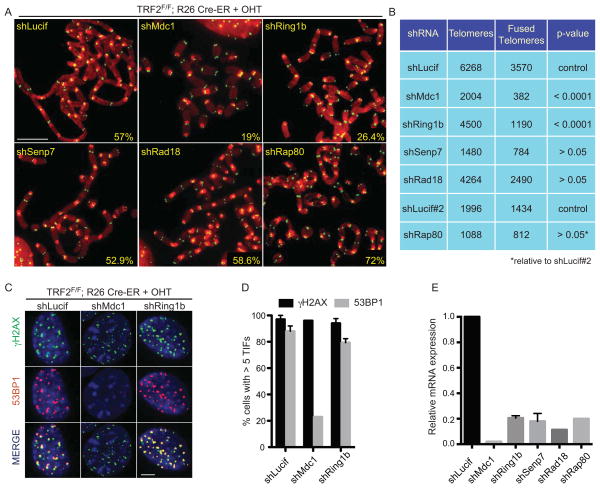
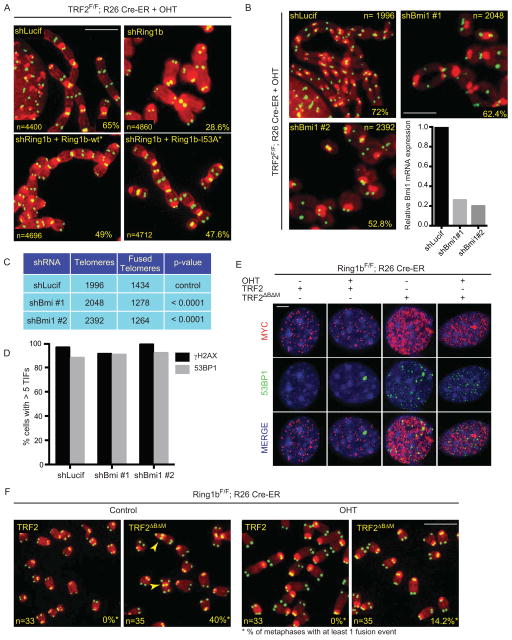
References
-
- Artandi SE, Chang S, Lee SL, Alson S, Gottlieb GJ, Chin L, DePinho RA. Telomere dysfunction promotes non-reciprocal translocations and epithelial cancers in mice. Nature. 2000;406:641–645. - PubMed
-
- Augereau A, T’Kint de Roodenbeke C, Simonet T, Bauwens S, Horard B, Callanan M, Leroux D, Jallades L, Salles G, Gilson E, et al. Telomeric damage in early stage of chronic lymphocytic leukemia correlates with shelterin dysregulation. Blood. 2011;118:1316–1322. - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
