

Hematopoietic not systemic impairment of Roquin expression accounts for intestinal inflammation in Roquin-deficient mice
- PMID: 24815331
- PMCID: PMC4017215
- DOI: 10.1038/srep04920
Hematopoietic not systemic impairment of Roquin expression accounts for intestinal inflammation in Roquin-deficient mice
Abstract
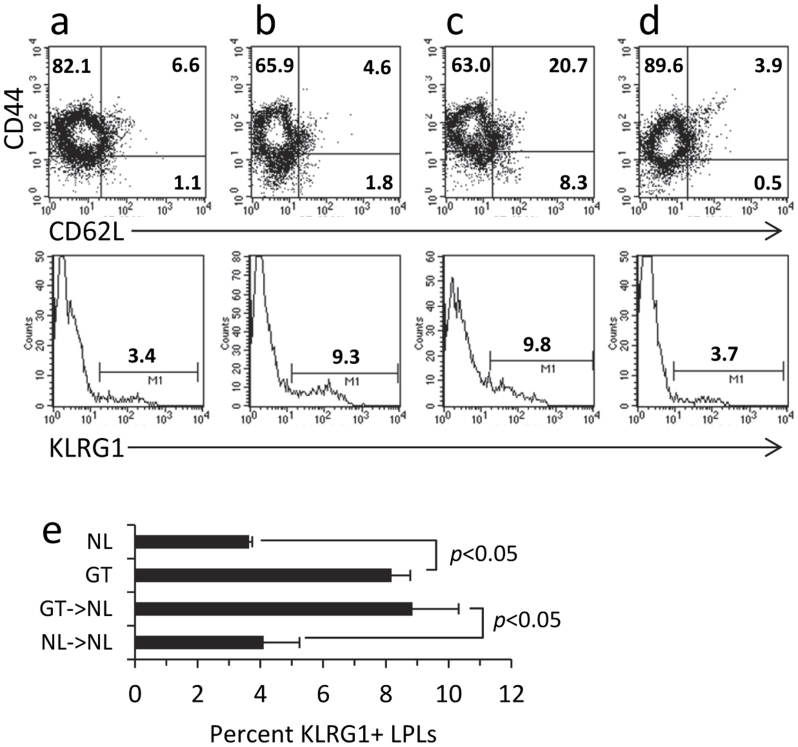
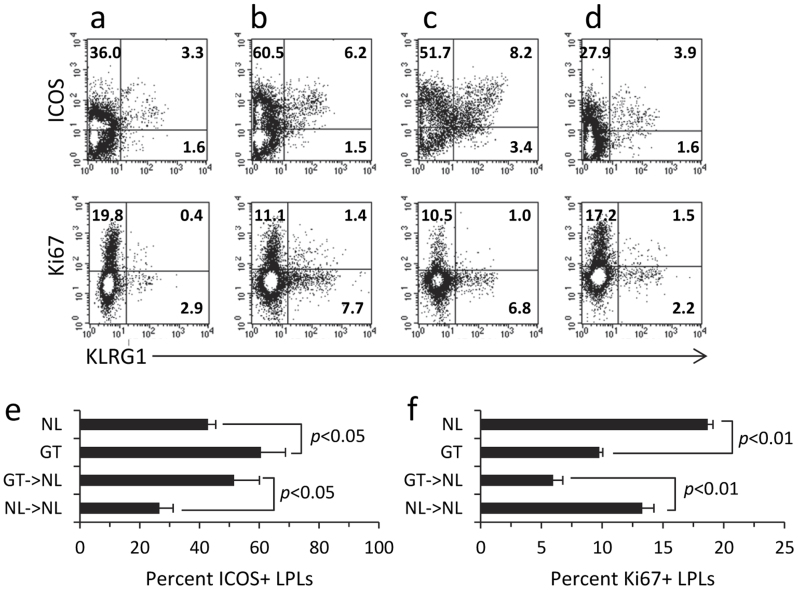
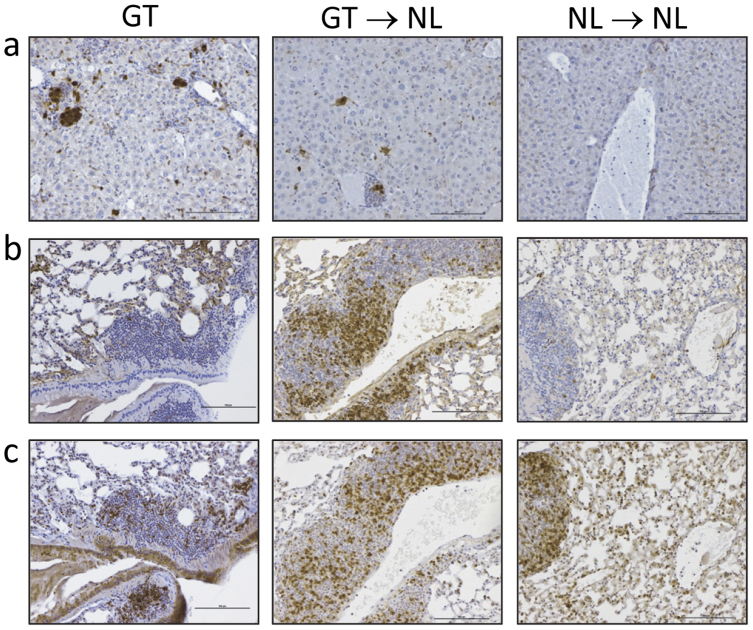
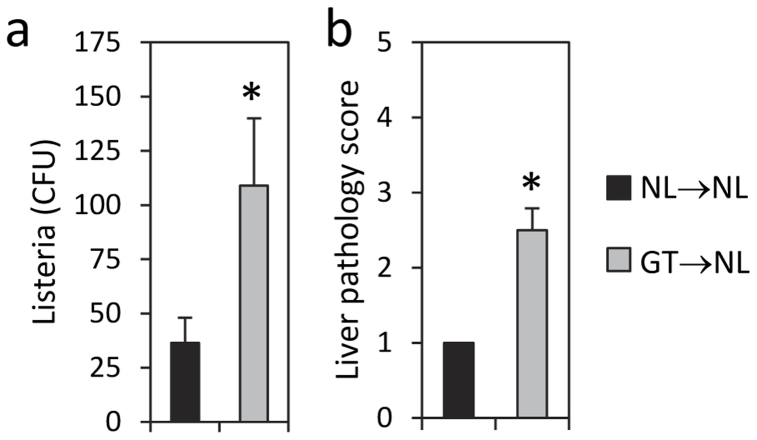
Roquin, an E3 ligase, is involved in curtailing autoimmune pathology as seen from studies using mice with mutated (Rc3h1(san/san)) or disrupted (Rc3h1(gt/gt)) Rc3h1 gene. The extent to which intestinal immunopathology is caused by insufficient Roquin expression in the immune system, or by Roquin impairment in non-hematopoietic cells, has not been determined. Using bone marrow cells from Rc3h1(gt/gt) mice transferred into irradiated normal mice (Rc3h1(gt/gt) → NL chimeras), we show that inflammation developed in the small intestine, kidney, lung, liver, and spleen. Proinflammatory cytokine levels were elevated in lamina propria lymphocytes (LPLs). Inflammation in the liver was accompanied by areas of hepatocyte apoptosis. Lung inflammation consisted of an influx of both T cells and B cells. Small intestinal LPLs had increased numbers of CD44(hi), CD62L(lo), KLRG1(+), ICOS(+) short-lived effector cells, indicating an influx of activated T cells. Following oral infection with L. monocytogenes, Rc3h1(gt/gt) → NL chimeras had more liver pathology and greater numbers of bacteria in the Peyer's patches than NL → NL chimeras. These findings demonstrate that small intestinal inflammation in Rc3h1(san/san) and Rc3h1(gt/gt) mice is due to a failure of Roquin expression in the immune system and not to insufficient systemic Roquin expression.
Figures



References
-
- Lin A. E. & Mak T. W. The role of E3 ligases in autoimmunity and the regulation of autoreactive T cells. Curr Opin Immunol 19, 665–673 (2007). - PubMed
-
- Vinuesa C. G. et al. A RING-type ubiquitin ligase family member required to repress follicular helper T cells and autoimmunity. Nature 435, 452–458 (2005). - PubMed
-
- Yu D. et al. Roquin represses autoimmunity by limiting inducible T-cell co-stimulator messenger RNA. Nature 450, 299–303 (2007). - PubMed
-
- Heissmeyer V. & Vogel K. U. Molecular control of Tfh-cell differentiation by Roquin family proteins. Immunol Rev 253, 273–289 (2013). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
