

Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity
- PMID: 24821781
- PMCID: PMC4078846
- DOI: 10.1073/pnas.1406974111
Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity
Abstract
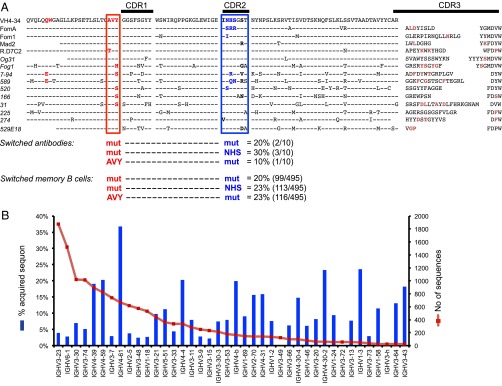
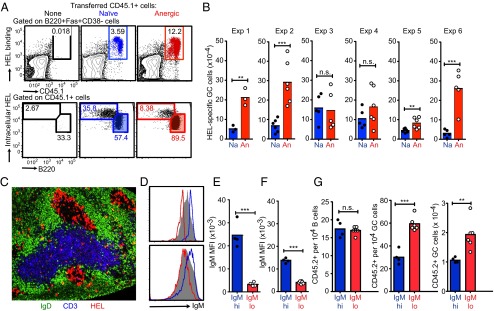
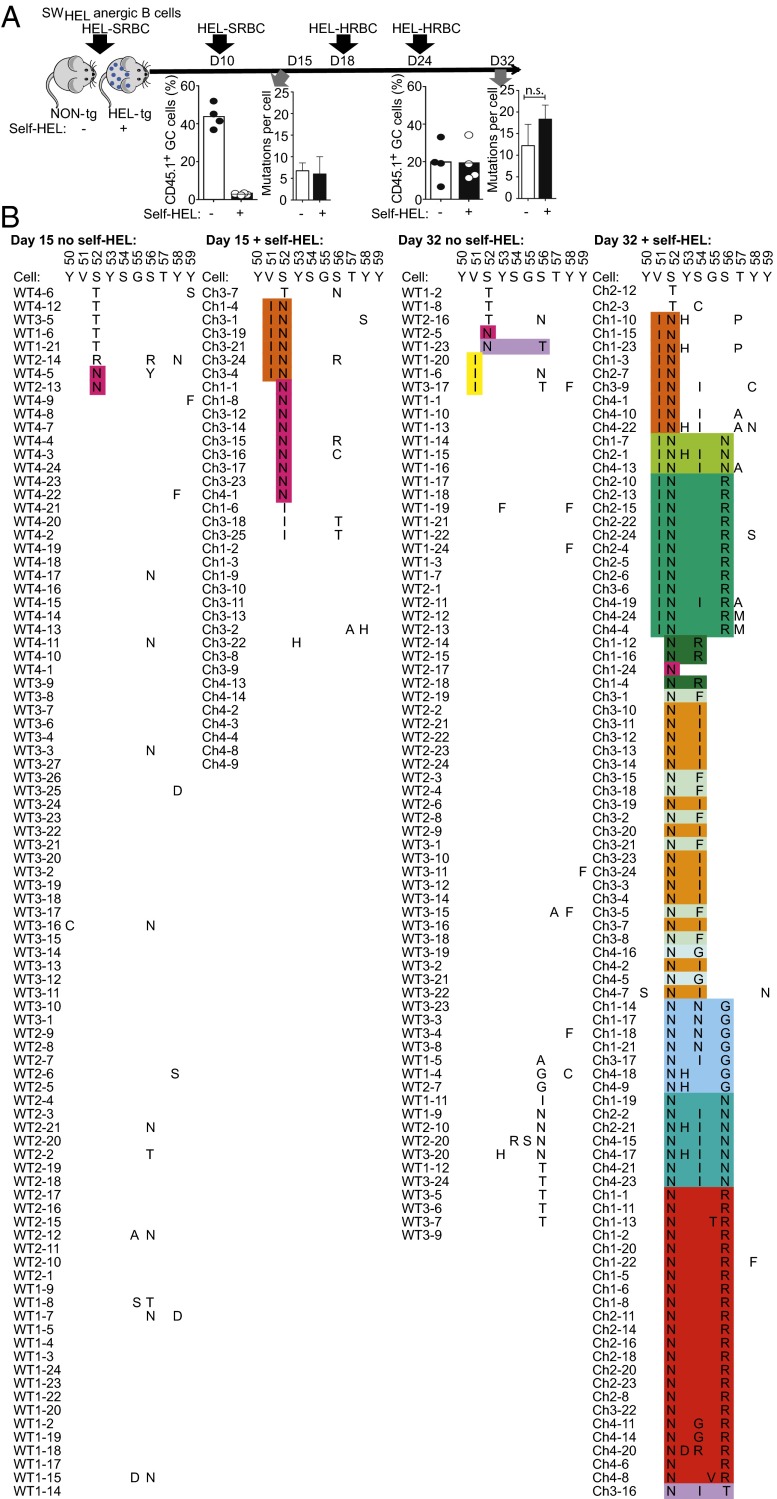
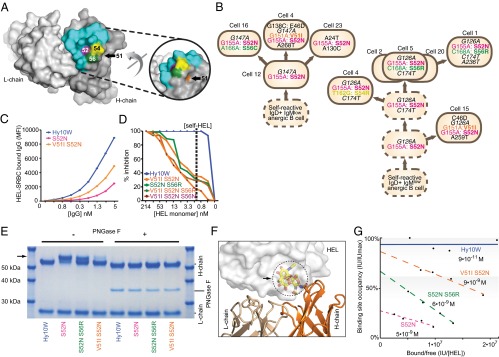
The best-understood mechanisms for achieving antibody self/non-self discrimination discard self-reactive antibodies before they can be tested for binding microbial antigens, potentially creating holes in the repertoire. Here we provide evidence for a complementary mechanism: retaining autoantibodies in the repertoire displayed as low levels of IgM and high IgD on anergic B cells, masking a varying proportion of autoantibody-binding sites with carbohydrates, and removing their self-reactivity by somatic hypermutation and selection in germinal centers (GCs). Analysis of human antibody sequences by deep sequencing of isotype-switched memory B cells or in IgG antibodies elicited against allogeneic RhD+ erythrocytes, vaccinia virus, rotavirus, or tetanus toxoid provides evidence for reactivation of anergic IgM(low) IgD+ IGHV4-34+ B cells and removal of cold agglutinin self-reactivity by hypermutation, often accompanied by mutations that inactivated an N-linked glycosylation sequon in complementarity-determining region 2 (CDR2). In a Hy10 antibody transgenic model where anergic B cells respond to a biophysically defined lysozyme epitope displayed on both foreign and self-antigens, cell transfers revealed that anergic IgM(low) IgD+ B cells form twice as many GC progeny as naïve IgM(hi) IgD+ counterparts. Their GC progeny were rapidly selected for CDR2 mutations that blocked 72% of antigen-binding sites with N-linked glycan, decreased affinity 100-fold, and then cleared the binding sites of blocking glycan. These results provide evidence for a mechanism to acquire self/non-self discrimination by somatic mutation away from self-reactivity, and reveal how varying the efficiency of N-glycosylation provides a mechanism to modulate antibody avidity.
Keywords: affinity maturation; autoimmunity; clonal selection; self-tolerance.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Redemption of autoreactive B cells.Proc Natl Acad Sci U S A. 2014 Jun 24;111(25):9022-3. doi: 10.1073/pnas.1407877111. Epub 2014 Jun 11. Proc Natl Acad Sci U S A. 2014. PMID: 24920593 Free PMC article. No abstract available.
References
-
- Burnet FM. The Clonal Selection Theory of Acquired Immunity. Cambridge, UK: Cambridge Univ Press; 1959.
-
- Goodnow CC, Ohashi PS. Immunological tolerance. In: Paul WE, editor. Fundamental Immunology. 7th Ed. Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins; 2013. pp. 765–794.
-
- Jerne NK. The somatic generation of immune recognition. Eur J Immunol. 1971;1(1):1–9. - PubMed
-
- Diaz M, Klinman NR. Relative roles of somatic and Darwinian evolution in shaping the antibody response. Immunol Res. 2000;21(2-3):89–102. - PubMed
-
- Goodnow CC, Crosbie J, Jorgensen H, Brink RA, Basten A. Induction of self-tolerance in mature peripheral B lymphocytes. Nature. 1989;342(6248):385–391. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
