

RIP1 suppresses innate immune necrotic as well as apoptotic cell death during mammalian parturition
- PMID: 24821786
- PMCID: PMC4040608
- DOI: 10.1073/pnas.1401857111
RIP1 suppresses innate immune necrotic as well as apoptotic cell death during mammalian parturition
Abstract
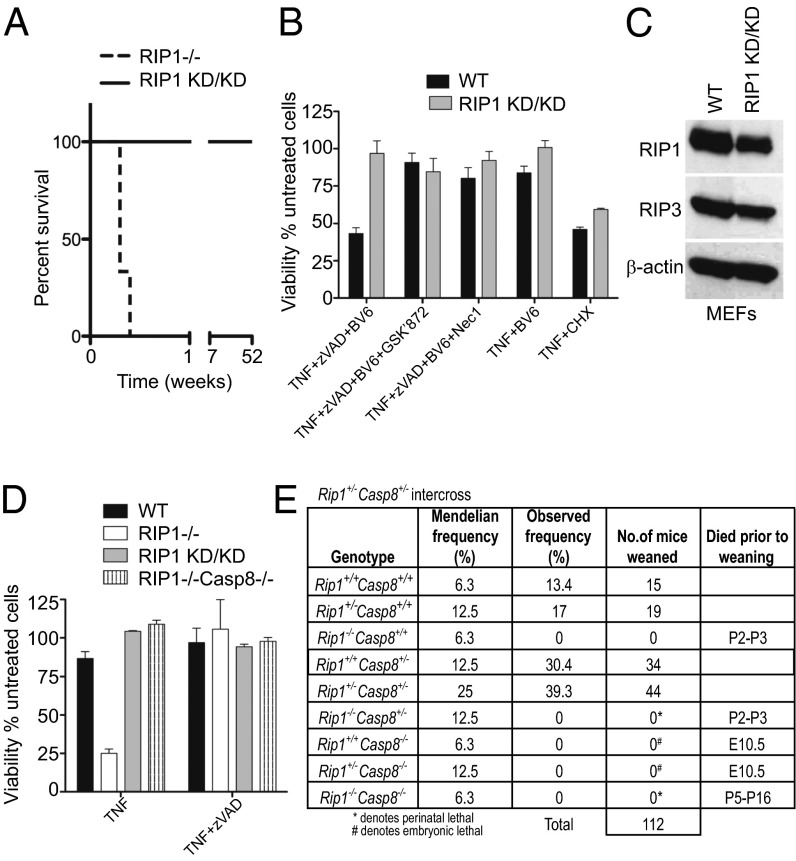
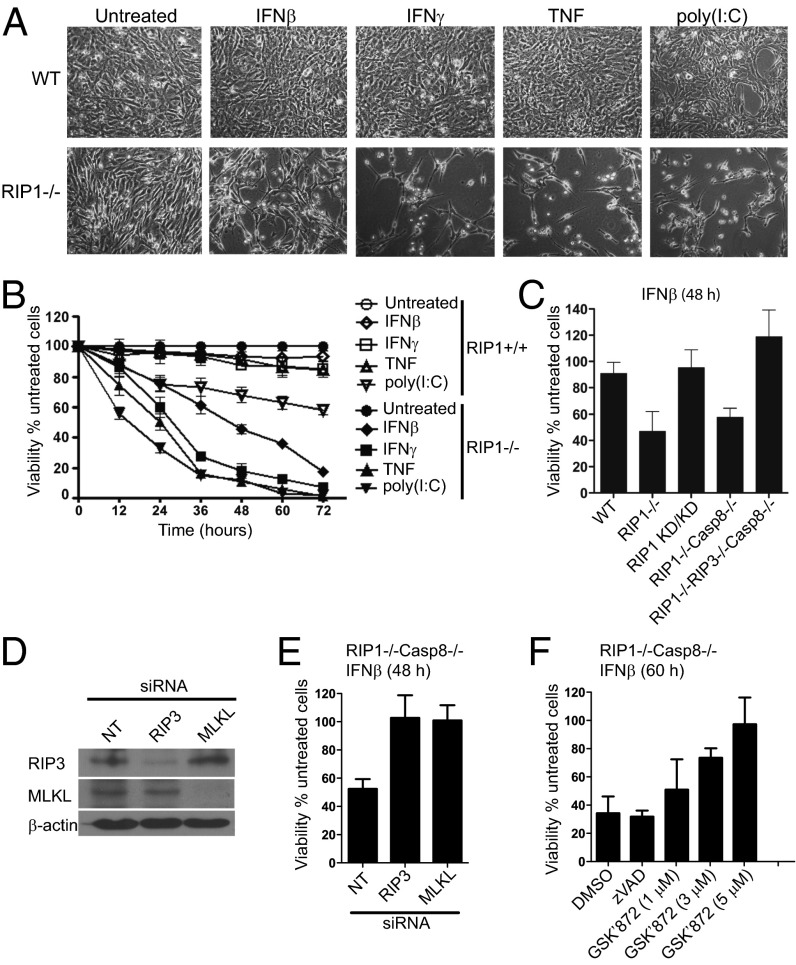
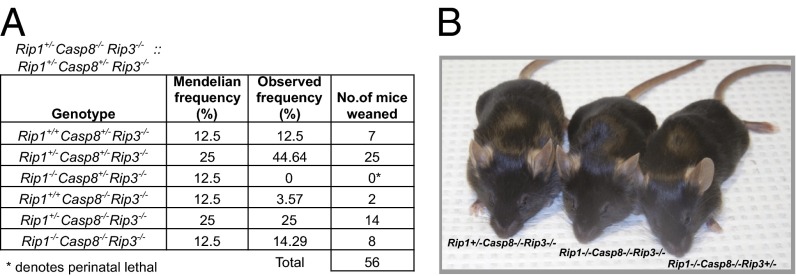
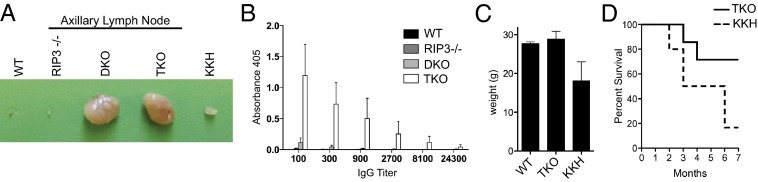
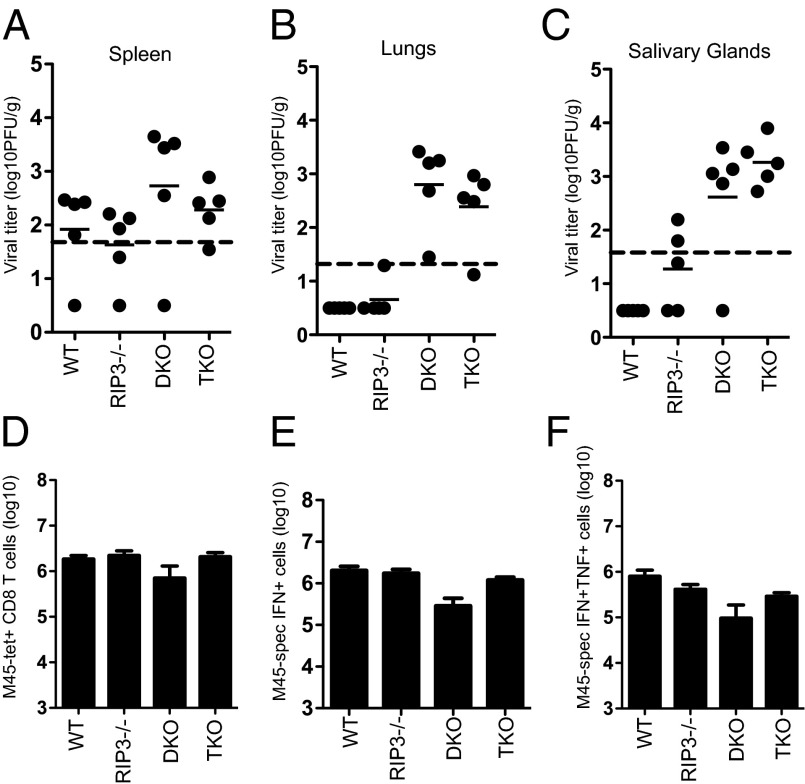
The pronecrotic kinase, receptor interacting protein (RIP1, also called RIPK1) mediates programmed necrosis and, together with its partner, RIP3 (RIPK3), drives midgestational death of caspase 8 (Casp8)-deficient embryos. RIP1 controls a second vital step in mammalian development immediately after birth, the mechanism of which remains unresolved. Rip1(-/-) mice display perinatal lethality, accompanied by gross immune system abnormalities. Here we show that RIP1 K45A (kinase dead) knockin mice develop normally into adulthood, indicating that development does not require RIP1 kinase activity. In the face of complete RIP1 deficiency, cells develop sensitivity to RIP3-mixed lineage kinase domain-like-mediated necroptosis as well as to Casp8-mediated apoptosis activated by diverse innate immune stimuli (e.g., TNF, IFN, double-stranded RNA). When either RIP3 or Casp8 is disrupted in combination with RIP1, the resulting double knockout mice exhibit slightly prolonged survival over RIP1-deficient animals. Surprisingly, triple knockout mice with combined RIP1, RIP3, and Casp8 deficiency develop into viable and fertile adults, with the capacity to produce normal levels of myeloid and lymphoid lineage cells. Despite the combined deficiency, these mice sustain a functional immune system that responds robustly to viral challenge. A single allele of Rip3 is tolerated in Rip1(-/-)Casp8(-/-)Rip3(+/-) mice, contrasting the need to eliminate both alleles of either Rip1 or Rip3 to rescue midgestational death of Casp8-deficient mice. These observations reveal a vital kinase-independent role for RIP1 in preventing pronecrotic as well as proapoptotic signaling events associated with life-threatening innate immune activation at the time of mammalian parturition.
Keywords: MLKL; herpesvirus; interferon.
Conflict of interest statement
Conflict of interest statement: P.J.G., J.B., and S.B.B. are employees of GlaxoSmithKline.
Figures
References
-
- Feoktistova M, Geserick P, Panayotova-Dimitrova D, Leverkus M. Pick your poison: The Ripoptosome, a cell death platform regulating apoptosis and necroptosis. Cell Cycle. 2012;11(3):460–467. - PubMed
-
- Kelliher MA, et al. The death domain kinase RIP mediates the TNF-induced NF-kappaB signal. Immunity. 1998;8(3):297–303. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
