

Partial sleep in the context of augmentation of brain function
- PMID: 24822040
- PMCID: PMC4013465
- DOI: 10.3389/fnsys.2014.00075
Partial sleep in the context of augmentation of brain function
Abstract
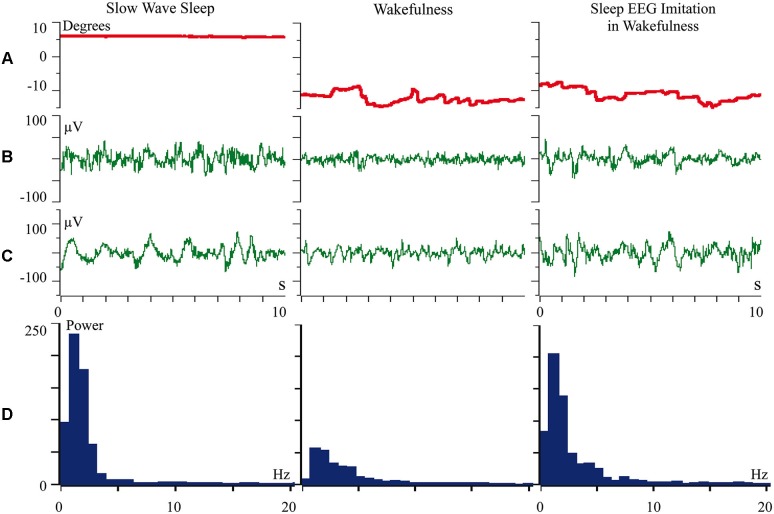
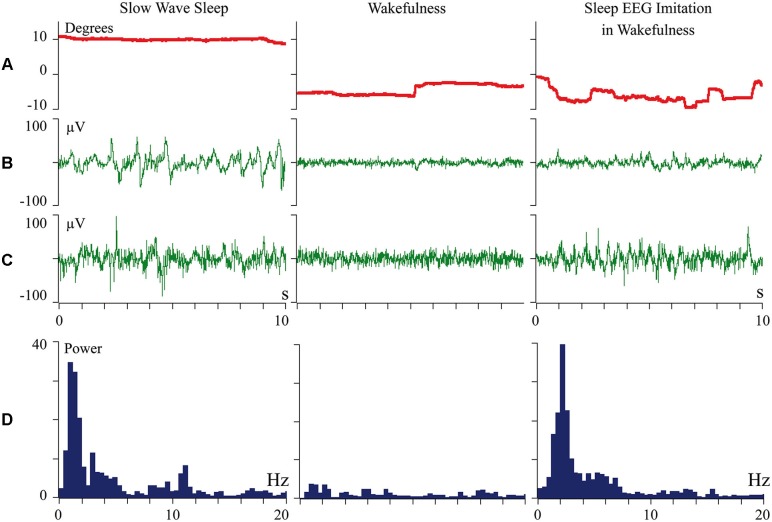
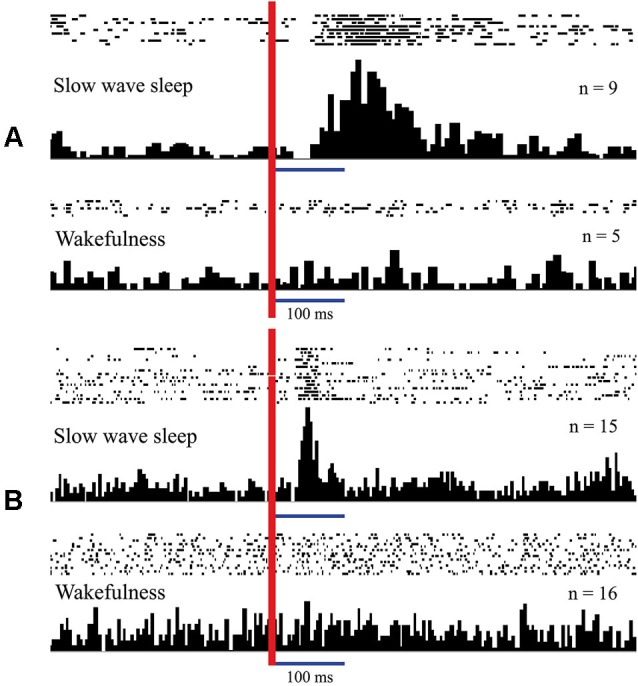
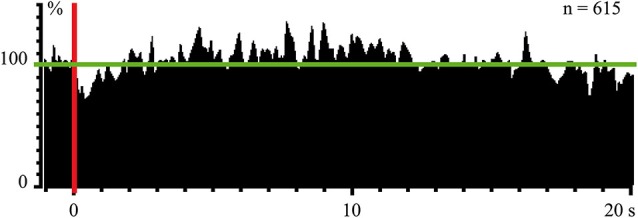
Inability to solve complex problems or errors in decision making is often attributed to poor brain processing, and raises the issue of brain augmentation. Investigation of neuronal activity in the cerebral cortex in the sleep-wake cycle offers insights into the mechanisms underlying the reduction in mental abilities for complex problem solving. Some cortical areas may transit into a sleep state while an organism is still awake. Such local sleep would reduce behavioral ability in the tasks for which the sleeping areas are crucial. The studies of this phenomenon have indicated that local sleep develops in high order cortical areas. This is why complex problem solving is mostly affected by local sleep, and prevention of local sleep might be a potential way of augmentation of brain function. For this approach to brain augmentation not to entail negative consequences for the organism, it is necessary to understand the functional role of sleep. Our studies have given an unexpected answer to this question. It was shown that cortical areas that process signals from extero- and proprioreceptors during wakefulness, switch to the processing of interoceptive information during sleep. It became clear that during sleep all "computational power" of the brain is directed to the restoration of the vital functions of internal organs. These results explain the logic behind the initiation of total and local sleep. Indeed, a mismatch between the current parameters of any visceral system and the genetically determined normal range would provide the feeling of tiredness, or sleep pressure. If an environmental situation allows falling asleep, the organism would transit to a normal total sleep in all cortical areas. However, if it is impossible to go to sleep immediately, partial sleep may develop in some cortical areas in the still behaviorally awake organism. This local sleep may reduce both the "intellectual power" and the restorative function of sleep for visceral organs.
Keywords: cerebral cortex; local sleep; sleep function; slow wave sleep; visceral control.
Figures
Similar articles
-
The state of sleep and the current brain paradigm.Front Syst Neurosci. 2015 Oct 13;9:139. doi: 10.3389/fnsys.2015.00139. eCollection 2015. Front Syst Neurosci. 2015. PMID: 26528146 Free PMC article.
-
[Sleep, emotions and the visceral control].Fiziol Cheloveka. 2013 Nov-Dec;39(6):31-44. Fiziol Cheloveka. 2013. PMID: 25509170 Review. Russian.
-
[The history of observations and some methodological features of the studies on local sleep].Zh Nevrol Psikhiatr Im S S Korsakova. 2020;120(9. Vyp. 2):91-97. doi: 10.17116/jnevro202012009291. Zh Nevrol Psikhiatr Im S S Korsakova. 2020. PMID: 33076652 Russian.
-
[Selective stimulations and lesions of the rat brain nuclei as the models for research of the human sleep pathology mechanisms].Glas Srp Akad Nauka Med. 2011;(51):85-97. Glas Srp Akad Nauka Med. 2011. PMID: 22165729 Review. Serbian.
-
Somatovisceral Convergence in Sleep-Wake Cycle: Transmitting Different Types of Information via the Same Pathway.Front Netw Physiol. 2022 Mar 2;2:840565. doi: 10.3389/fnetp.2022.840565. eCollection 2022. Front Netw Physiol. 2022. PMID: 36926092 Free PMC article.
Cited by
-
Therapeutic Effects of Electrical Stimulation: Interpretations and Predictions Based on the Visceral Theory of Sleep.Front Neurosci. 2018 Feb 12;12:65. doi: 10.3389/fnins.2018.00065. eCollection 2018. Front Neurosci. 2018. PMID: 29483861 Free PMC article. No abstract available.
-
Alteration of coupling between brain and heart induced by sedation with propofol and midazolam.PLoS One. 2019 Jul 17;14(7):e0219238. doi: 10.1371/journal.pone.0219238. eCollection 2019. PLoS One. 2019. PMID: 31314775 Free PMC article.
-
Sleep in Normal Aging, Homeostatic and Circadian Regulation and Vulnerability to Sleep Deprivation.Brain Sci. 2021 Jul 29;11(8):1003. doi: 10.3390/brainsci11081003. Brain Sci. 2021. PMID: 34439622 Free PMC article. Review.
-
The state of sleep and the current brain paradigm.Front Syst Neurosci. 2015 Oct 13;9:139. doi: 10.3389/fnsys.2015.00139. eCollection 2015. Front Syst Neurosci. 2015. PMID: 26528146 Free PMC article.
-
The Mysterious Island: Insula and Its Dual Function in Sleep and Wakefulness.Front Syst Neurosci. 2021 Feb 11;14:592660. doi: 10.3389/fnsys.2020.592660. eCollection 2020. Front Syst Neurosci. 2021. PMID: 33643002 Free PMC article.
References
-
- Akeyson E. W., Schramm L. P. (1994). Splanchnic and somatic afferent convergence on cervical spinal neurons of the rat. Am. J. Physiol. 266(Suppl. 1), R268–R276 - PubMed
-
- Amassian V. E. (1951). Cortical representation of visceral afferents. J. Neurophysiol. 14, 435–446 - PubMed
-
- Bailey P., Bremer F. (1938). A sensory cortical representation of the vagus nerve. With a note on the effects of low blood pressure on the cortical electrogramm. J. Neurophysiol. 1, 405–414
LinkOut - more resources
Full Text Sources
Other Literature Sources