

Versatile microRNA biogenesis in animals and their viruses
- PMID: 24823351
- PMCID: PMC4156499
- DOI: 10.4161/rna.28985
Versatile microRNA biogenesis in animals and their viruses
Abstract
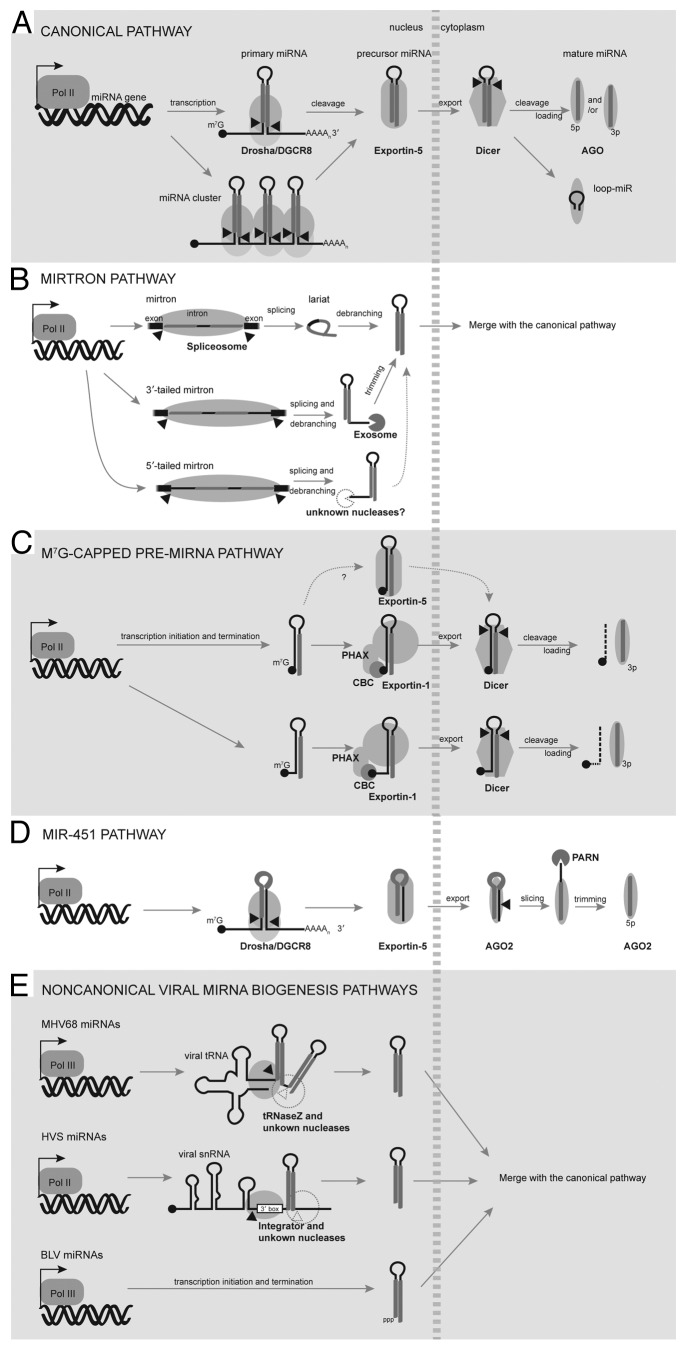
MicroRNAs (miRNAs) are ubiquitous gene regulators that modulate essential cellular processes at the post-transcriptional level. In metazoans and their viruses, most miRNAs are produced from hairpin-containing primary transcripts that are sequentially cleaved by nuclear Drosha and cytoplasmic Dicer. In the last decade, alternative mechanisms that bypass either the Drosha or Dicer cleavage step have emerged, increasing the complexity of the miRNA regulatory network. Here, we highlight non-canonical pathways that generate miRNAs using a variety of molecular machineries that play fundamental roles in the biogenesis and processing of other classes of cellular RNAs.
Keywords: AGO2 slicer activity; Dicer; Integrator; Microprocessor; exportin-5; m7G-capped pre-miRNA; microRNA biogenesis; mirtron; tRNaseZ; viral microRNA.
Figures
References
-
- Krol J, Loedige I, Filipowicz W. The widespread regulation of microRNA biogenesis, function and decay. Nat Rev Genet. 2010;11:597–610. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources