

Enzyme architecture: the effect of replacement and deletion mutations of loop 6 on catalysis by triosephosphate isomerase
- PMID: 24825099
- PMCID: PMC4051426
- DOI: 10.1021/bi500458t
Enzyme architecture: the effect of replacement and deletion mutations of loop 6 on catalysis by triosephosphate isomerase
Abstract
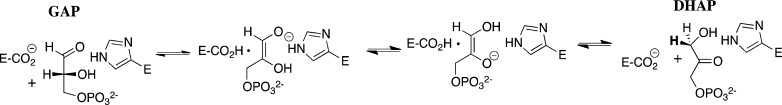
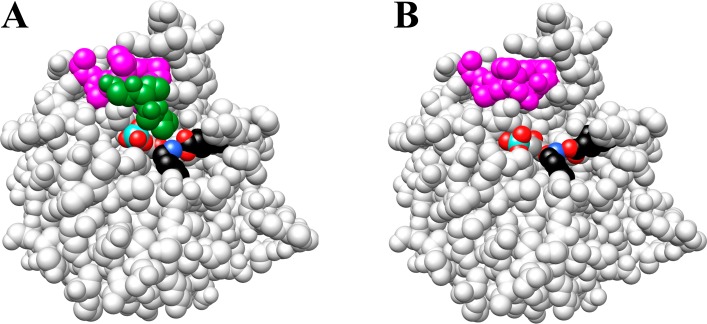
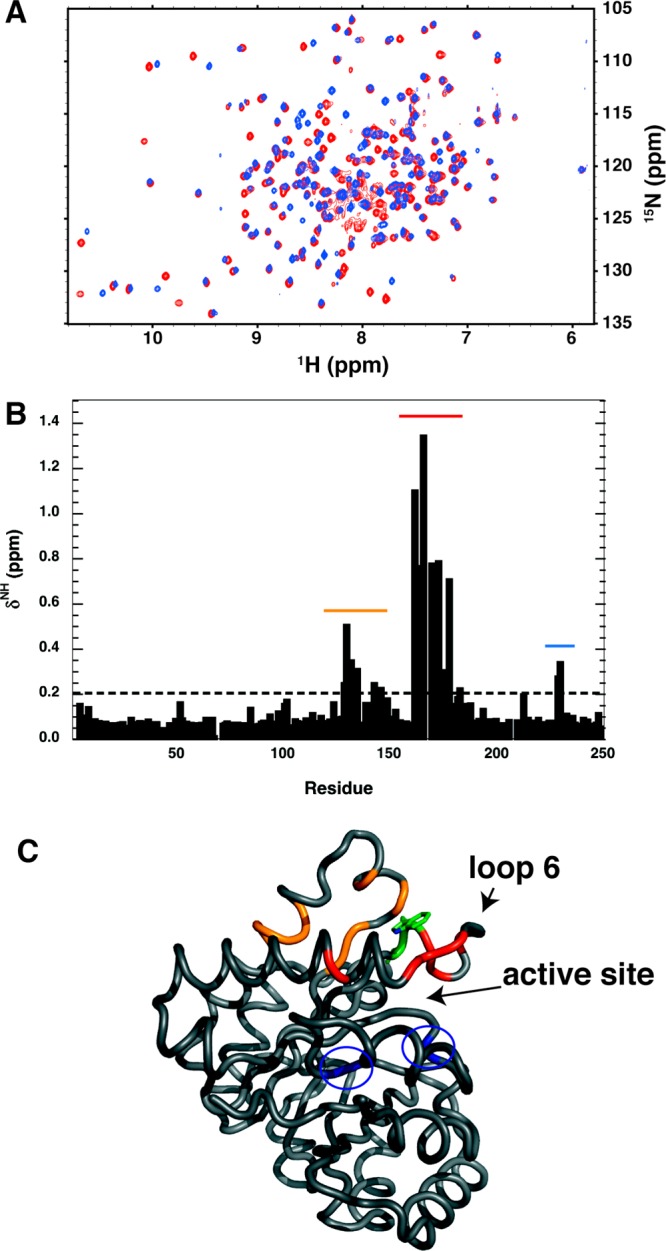
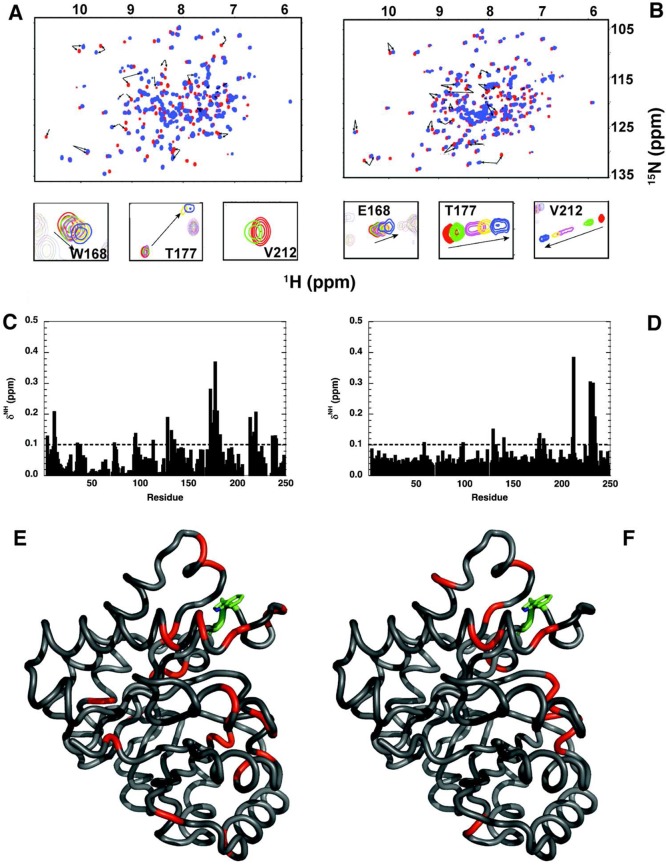
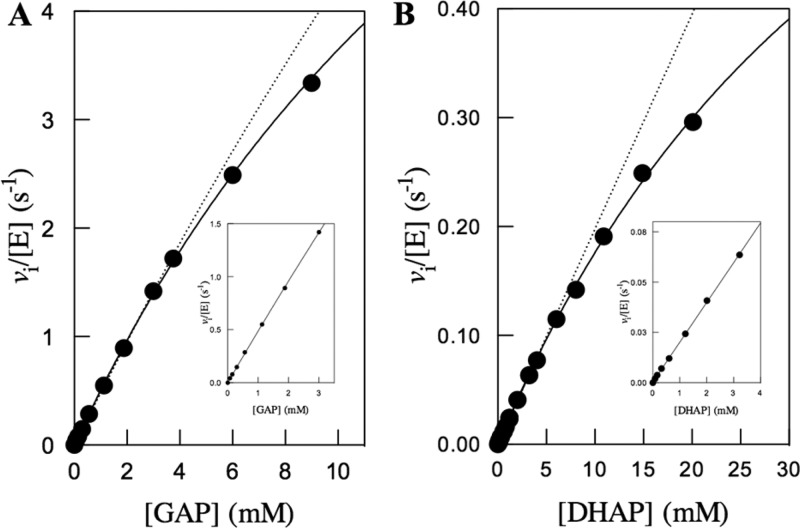
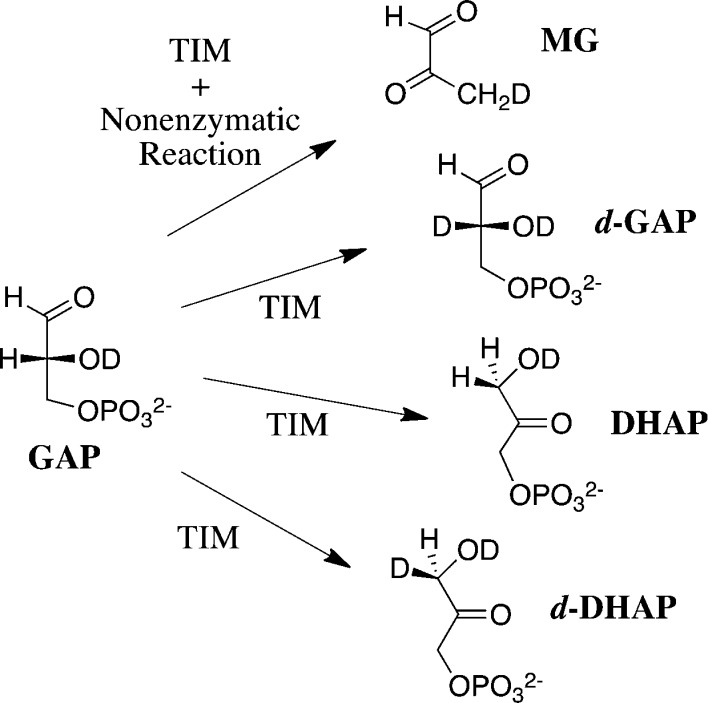
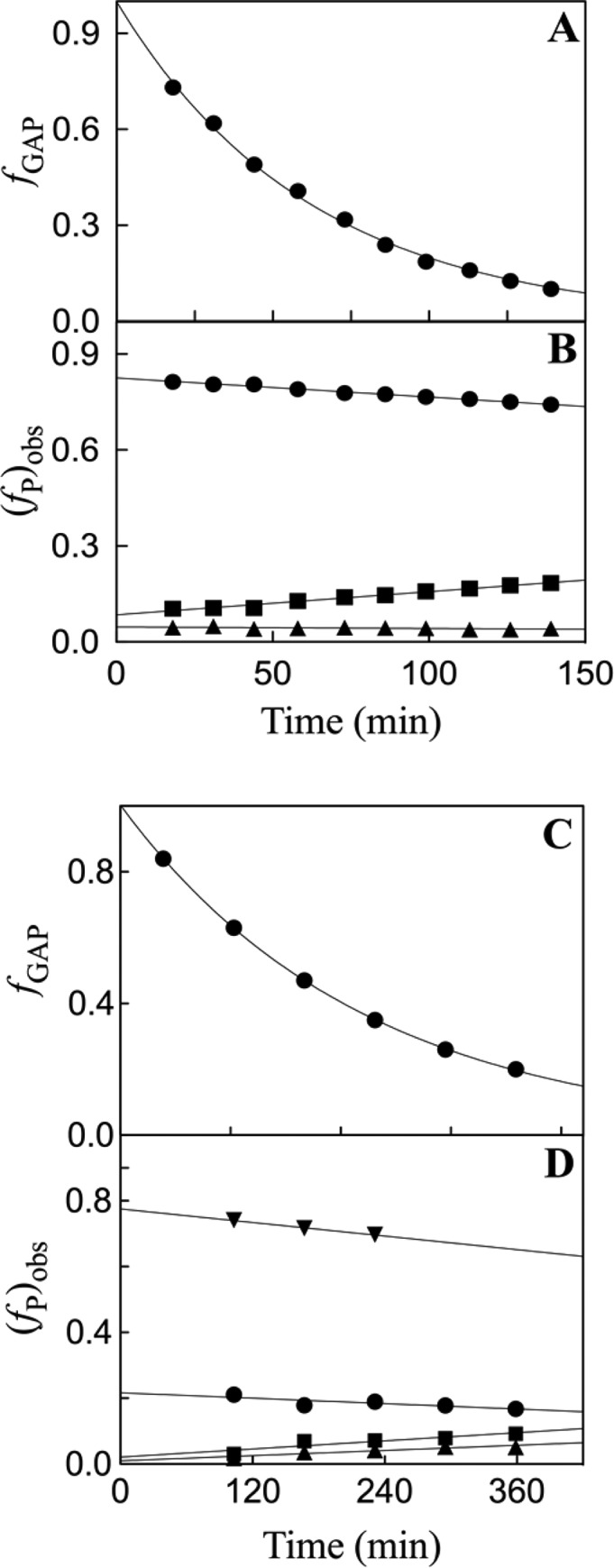
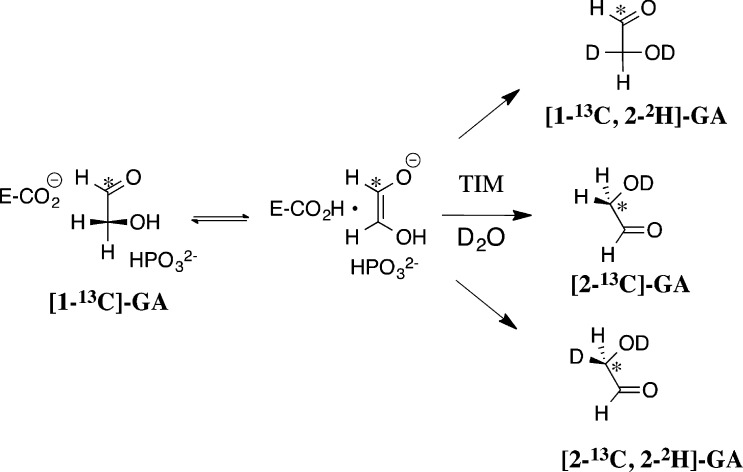
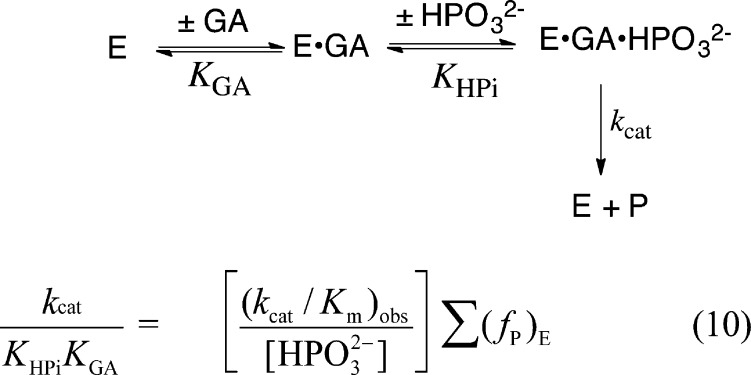
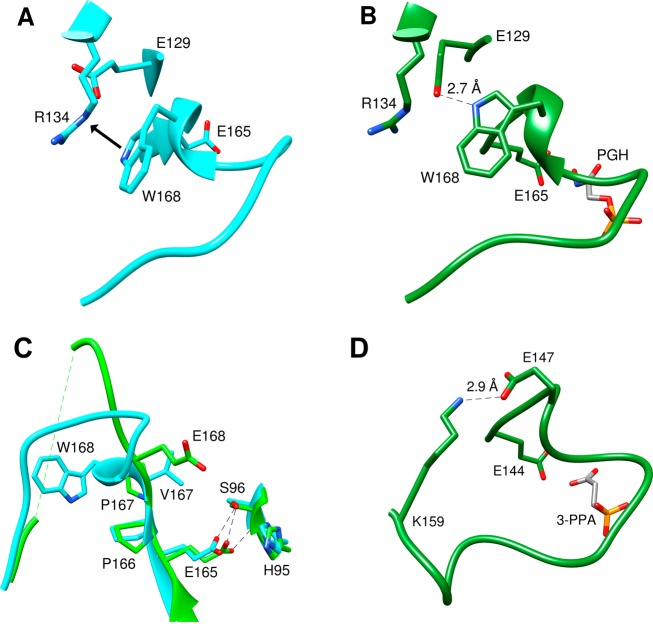
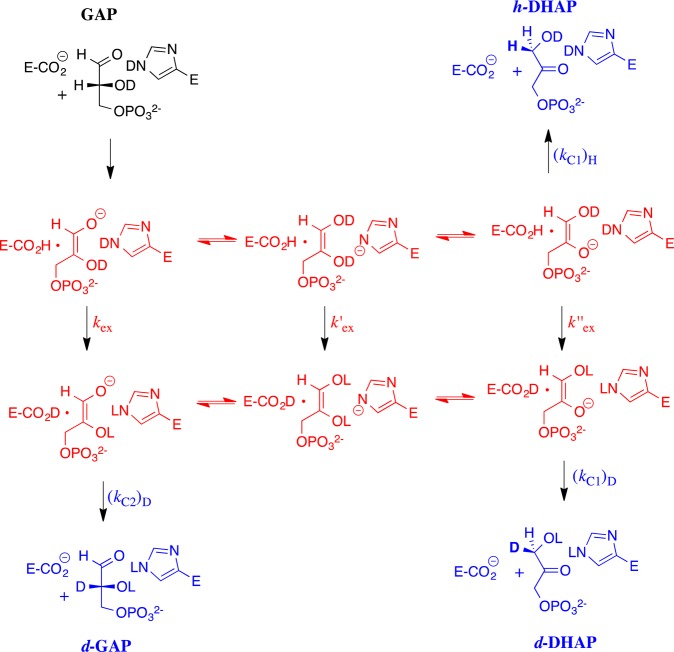
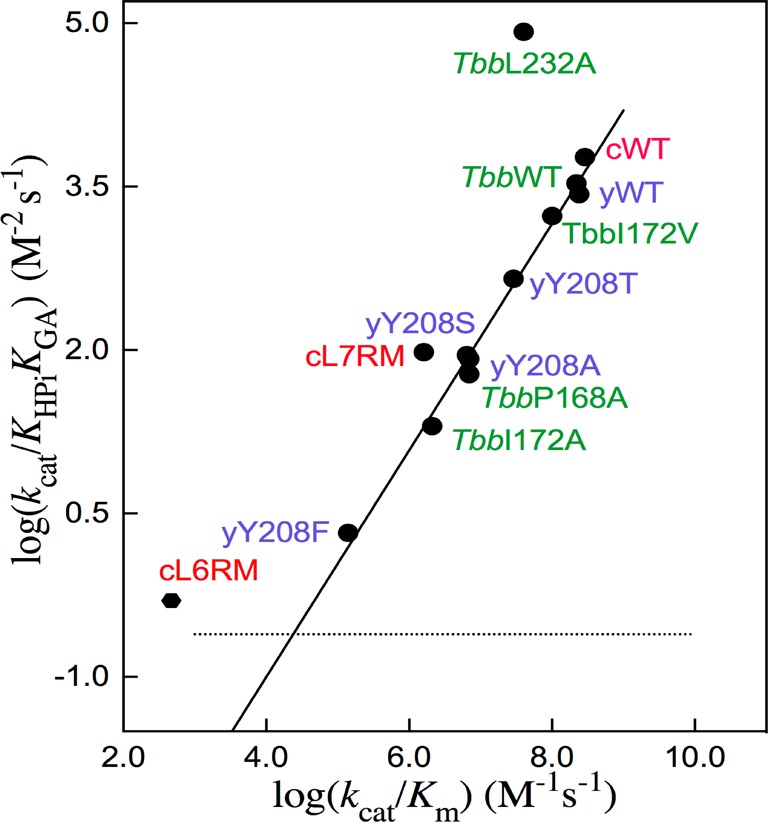
Two mutations of the phosphodianion gripper loop in chicken muscle triosephosphate isomerase (cTIM) were examined: (1) the loop deletion mutant (LDM) formed by removal of residues 170-173 [Pompliano, D. L., et al. (1990) Biochemistry 29, 3186-3194] and (2) the loop 6 replacement mutant (L6RM), in which the N-terminal hinge sequence of TIM from eukaryotes, 166-PXW-168 (X = L or V), is replaced by the sequence from archaea, 166-PPE-168. The X-ray crystal structure of the L6RM shows a large displacement of the side chain of E168 from that for W168 in wild-type cTIM. Solution nuclear magnetic resonance data show that the L6RM results in significant chemical shift changes in loop 6 and surrounding regions, and that the binding of glycerol 3-phosphate (G3P) results in chemical shift changes for nuclei at the active site of the L6RM that are smaller than those of wild-type cTIM. Interactions with loop 6 of the L6RM stabilize the enediolate intermediate toward the elimination reaction catalyzed by the LDM. The LDM and L6RM result in 800000- and 23000-fold decreases, respectively, in kcat/Km for isomerization of GAP. Saturation of the LDM, but not the L6RM, by substrate and inhibitor phosphoglycolate is detected by steady-state kinetic analyses. We propose, on the basis of a comparison of X-ray crystal structures for wild-type TIM and the L6RM, that ligands bind weakly to the L6RM because a large fraction of the ligand binding energy is utilized to overcome destabilizing electrostatic interactions between the side chains of E168 and E129 that are predicted to develop in the loop-closed enzyme. Similar normalized yields of DHAP, d-DHAP, and d-GAP are formed in LDM- and L6RM-catalyzed reactions of GAP in D2O. The smaller normalized 12-13% yield of DHAP and d-DHAP observed for the mutant cTIM-catalyzed reactions compared with the 79% yield of these products for wild-type cTIM suggests that these mutations impair the transfer of a proton from O-2 to O-1 at the initial enediolate phosphate intermediate. No products are detected for the LDM-catalyzed isomerization reactions in D2O of [1-(13)C]GA and HPi, but the L6RM-catalyzed reaction in the presence of 0.020 M dianion gives a 2% yield of the isomerization product [2-(13)C,2-(2)H]GA.
Figures


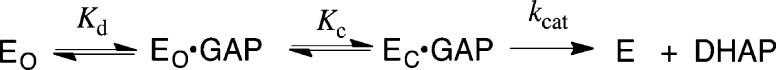
References
-
- Fothergill-Gilmore L. A. (1986) The evolution of the glycolytic pathway. Trends Biochem. Sci. 11, 47–51.
-
- Knowles J. R.; Albery W. J. (1977) Perfection in enzyme catalysis: The energetics of triosephosphate isomerase. Acc. Chem. Res. 10, 105–111.
-
- Knowles J. R. (1991) To build an enzyme. Philos. Trans. R. Soc. London, Ser. B 332, 115–121. - PubMed
-
- Waley S. G.; Miller J. C.; Rose I. A.; O’Connell E. L. (1970) Identification of site in triose phosphate isomerase labeled by glycidol phosphate. Nature 227, 181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
