

Stomatal and pavement cell density linked to leaf internal CO2 concentration
- PMID: 24825295
- PMCID: PMC4217638
- DOI: 10.1093/aob/mcu095
Stomatal and pavement cell density linked to leaf internal CO2 concentration
Abstract
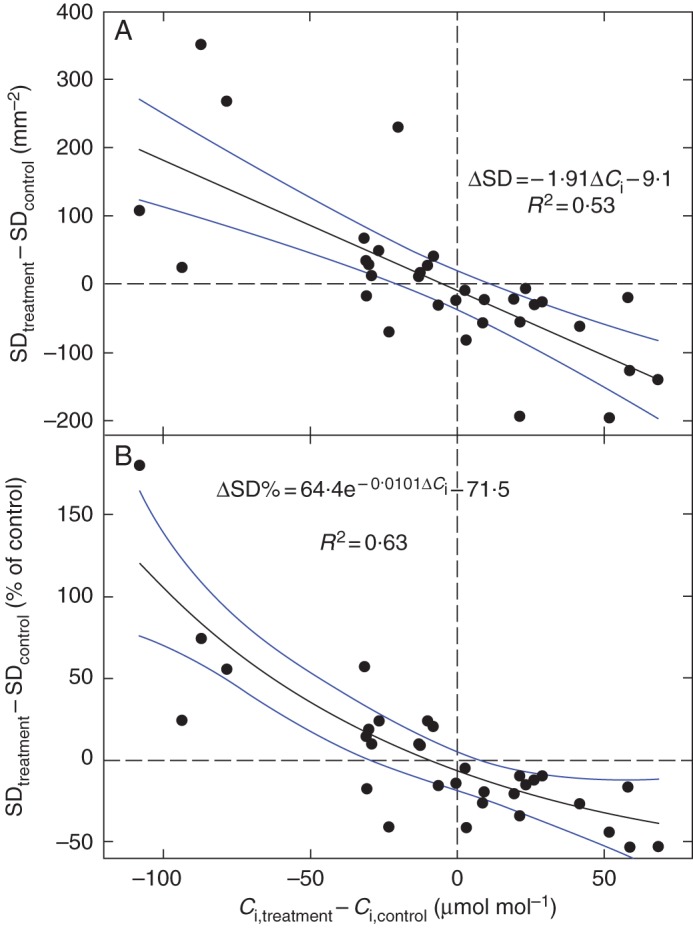
Background and aims: Stomatal density (SD) generally decreases with rising atmospheric CO2 concentration, Ca. However, SD is also affected by light, air humidity and drought, all under systemic signalling from older leaves. This makes our understanding of how Ca controls SD incomplete. This study tested the hypotheses that SD is affected by the internal CO2 concentration of the leaf, Ci, rather than Ca, and that cotyledons, as the first plant assimilation organs, lack the systemic signal.
Methods: Sunflower (Helianthus annuus), beech (Fagus sylvatica), arabidopsis (Arabidopsis thaliana) and garden cress (Lepidium sativum) were grown under contrasting environmental conditions that affected Ci while Ca was kept constant. The SD, pavement cell density (PCD) and stomatal index (SI) responses to Ci in cotyledons and the first leaves of garden cress were compared. (13)C abundance (δ(13)C) in leaf dry matter was used to estimate the effective Ci during leaf development. The SD was estimated from leaf imprints.
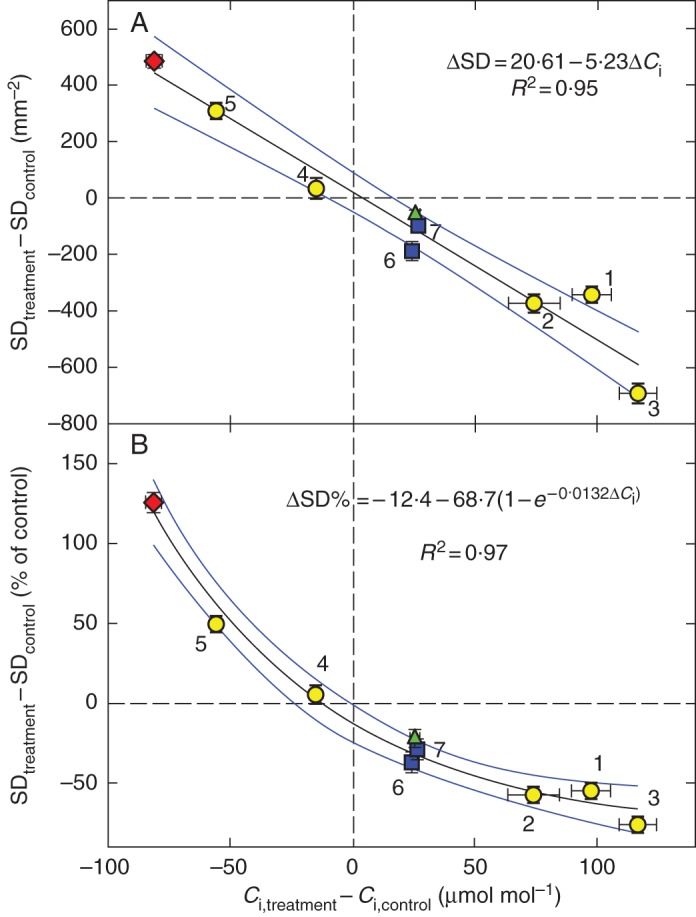
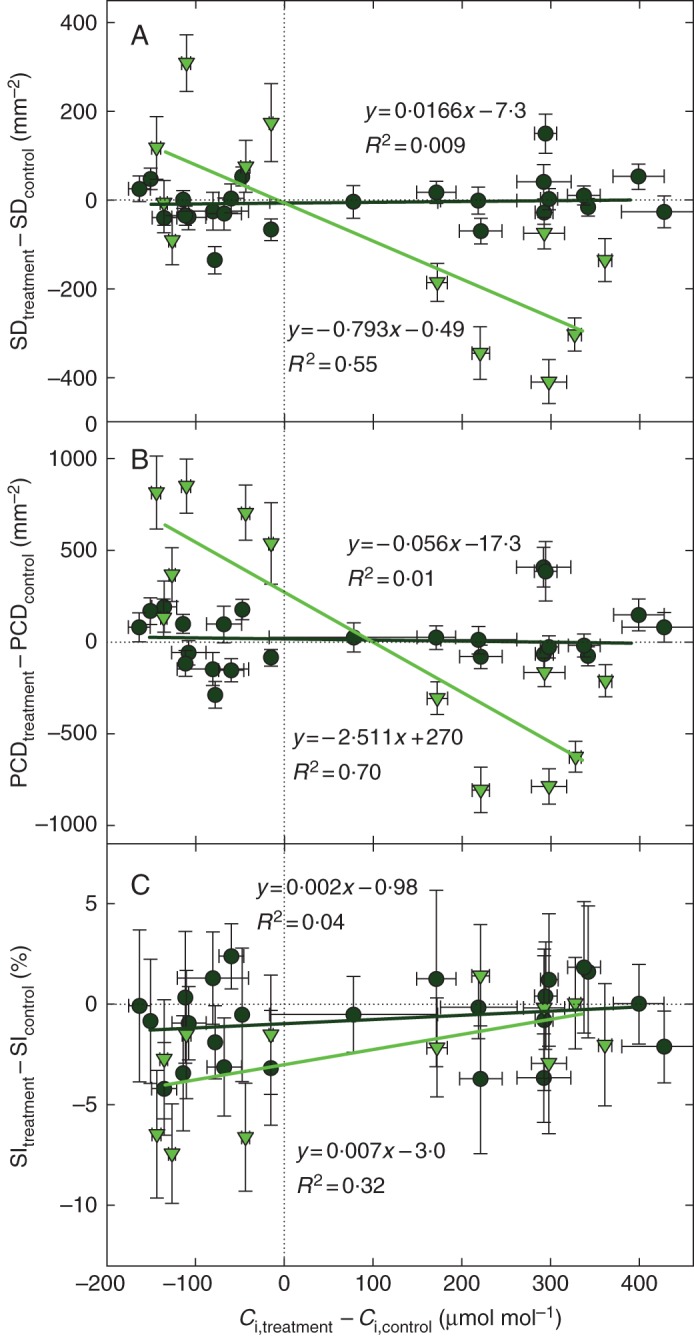
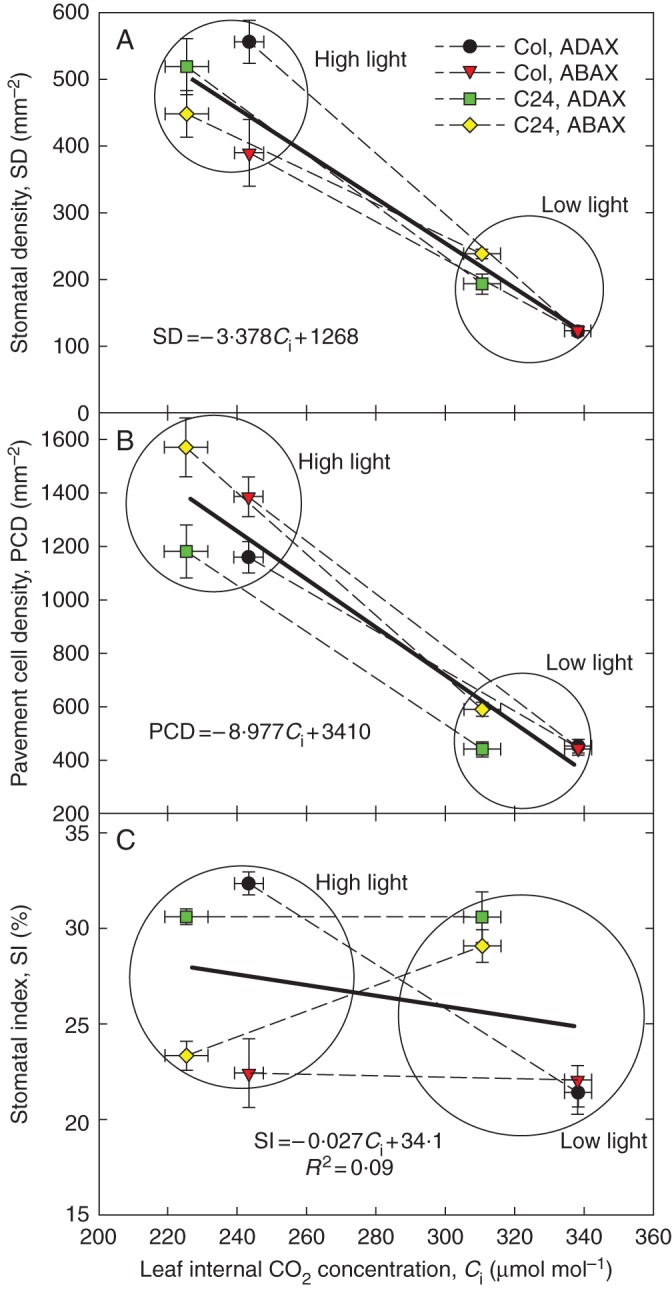
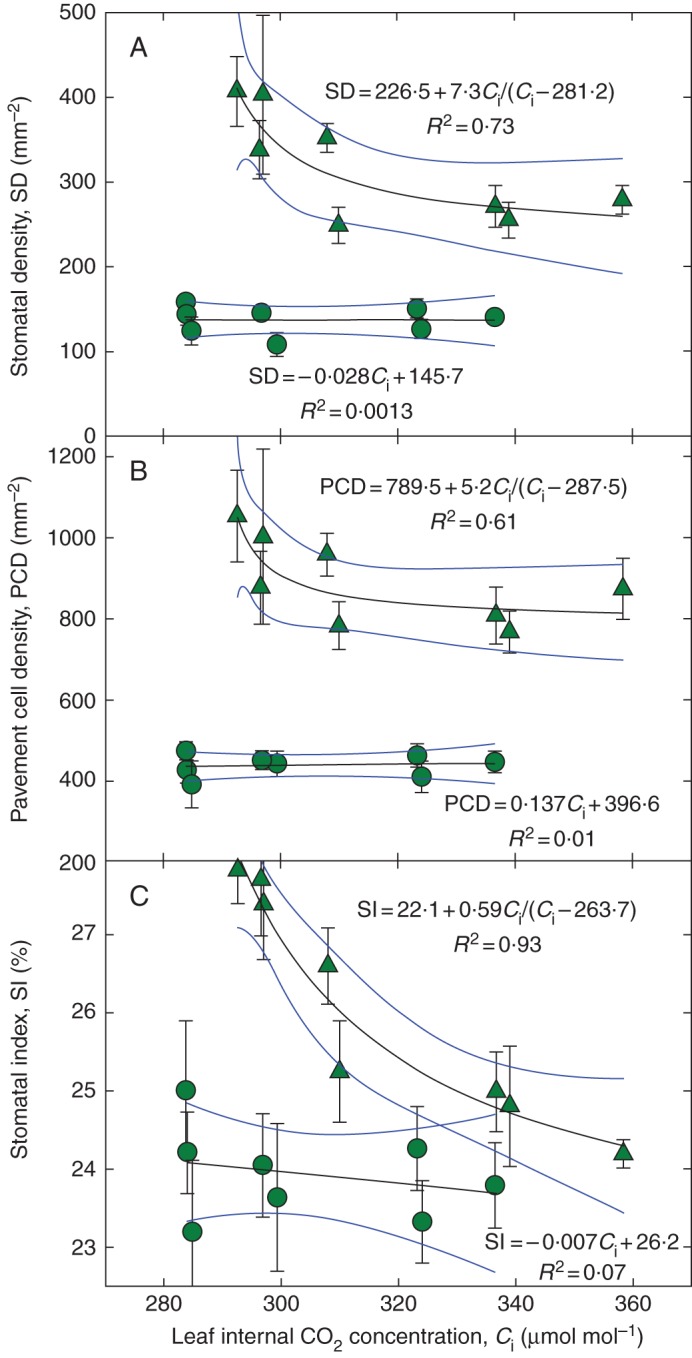
Key results: SD correlated negatively with Ci in leaves of all four species and under three different treatments (irradiance, abscisic acid and osmotic stress). PCD in arabidopsis and garden cress responded similarly, so that SI was largely unaffected. However, SD and PCD of cotyledons were insensitive to Ci, indicating an essential role for systemic signalling.
Conclusions: It is proposed that Ci or a Ci-linked factor plays an important role in modulating SD and PCD during epidermis development and leaf expansion. The absence of a Ci-SD relationship in the cotyledons of garden cress indicates the key role of lower-insertion CO2 assimilation organs in signal perception and its long-distance transport.
Keywords: 13C discrimination; Arabidopsis thaliana; Fagus sylvatica; Helianthus annuus; Lepidium sativum; Stomatal density; cotyledons; leaf internal CO2; pavement cells; stomata development.
© The Author 2014. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
Similar articles
-
Elevated CO2 promotes satellite stomata production in young cotyledons of Arabidopsis thaliana.Genes Cells. 2020 Jul;25(7):475-482. doi: 10.1111/gtc.12773. Epub 2020 May 12. Genes Cells. 2020. PMID: 32294311
-
Effects of elevated carbon dioxide on stomatal characteristics and carbon isotope ratio of Arabidopsis thaliana ecotypes originating from an altitudinal gradient.Physiol Plant. 2017 Jan;159(1):74-92. doi: 10.1111/ppl.12486. Epub 2016 Nov 6. Physiol Plant. 2017. PMID: 27514017
-
Natural variation in stomatal abundance of Arabidopsis thaliana includes cryptic diversity for different developmental processes.Ann Bot. 2011 Jun;107(8):1247-58. doi: 10.1093/aob/mcr060. Epub 2011 Mar 28. Ann Bot. 2011. PMID: 21447490 Free PMC article.
-
Abscisic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus-response analyses.Ann Bot. 2012 Jan;109(1):5-17. doi: 10.1093/aob/mcr252. Epub 2011 Oct 12. Ann Bot. 2012. PMID: 21994053 Free PMC article. Review.
-
The functional significance of the stomatal size to density relationship: Interaction with atmospheric [CO2] and role in plant physiological behaviour.Sci Total Environ. 2023 Mar 10;863:160908. doi: 10.1016/j.scitotenv.2022.160908. Epub 2022 Dec 16. Sci Total Environ. 2023. PMID: 36535478 Review.
Cited by
-
CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions.Trends Plant Sci. 2016 Jan;21(1):16-30. doi: 10.1016/j.tplants.2015.08.014. Epub 2015 Oct 5. Trends Plant Sci. 2016. PMID: 26482956 Free PMC article. Review.
-
Elevation-related variation in leaf stomatal traits as a function of plant functional type: evidence from Changbai Mountain, China.PLoS One. 2014 Dec 17;9(12):e115395. doi: 10.1371/journal.pone.0115395. eCollection 2014. PLoS One. 2014. PMID: 25517967 Free PMC article.
-
Stomatal design principles in synthetic and real leaves.J R Soc Interface. 2016 Nov;13(124):20160535. doi: 10.1098/rsif.2016.0535. J R Soc Interface. 2016. PMID: 27807270 Free PMC article.
-
Polycomb repressive complex 2 facilitates the transition from heterotrophy to photoautotrophy during seedling emergence.Plant Cell. 2025 Jul 1;37(7):koaf148. doi: 10.1093/plcell/koaf148. Plant Cell. 2025. PMID: 40515680 Free PMC article.
-
Integrating stomatal physiology and morphology: evolution of stomatal control and development of future crops.Oecologia. 2021 Dec;197(4):867-883. doi: 10.1007/s00442-021-04857-3. Epub 2021 Jan 30. Oecologia. 2021. PMID: 33515295 Free PMC article.
References
-
- Abrams MD. Genotypic and phenotypic variation as stress adaptations in temperate tree species – a review of several case-studies. Tree Physiology. 1994;14:833–842. - PubMed
-
- Ainsworth EA, Rogers A. The response of photosynthesis and stomatal conductance to rising CO2: mechanisms and environmental interactions. Plant, Cell and Environment. 2007;30:258–270. - PubMed
-
- Aucour AM, Gomez B, Sheppard SMF, Thevenard F. Delta C-13 and stomatal number variability in the Cretaceous conifer Frenelopsis. Palaeogeography Palaeoclimatology Palaeoecology. 2008;257:462–473.
-
- Bakker JC. Effects of humidity on stomatal density and its relation to leaf conductance. Scientia Horticulturae. 1991;48:205–212.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
