

Androgen-dependent sertoli cell tight junction remodeling is mediated by multiple tight junction components
- PMID: 24825397
- PMCID: PMC4075161
- DOI: 10.1210/me.2013-1134
Androgen-dependent sertoli cell tight junction remodeling is mediated by multiple tight junction components
Abstract
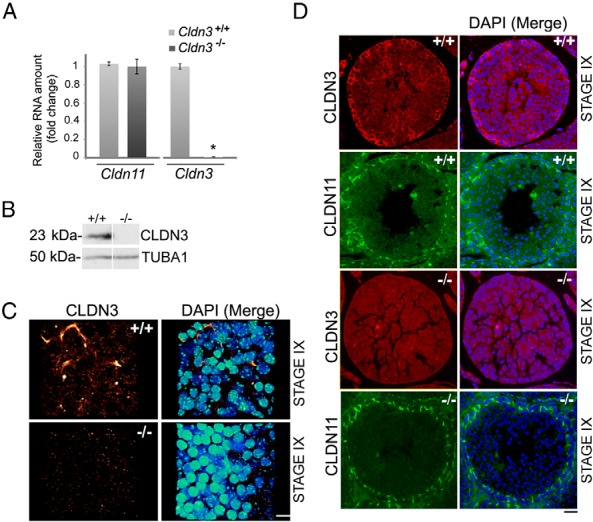
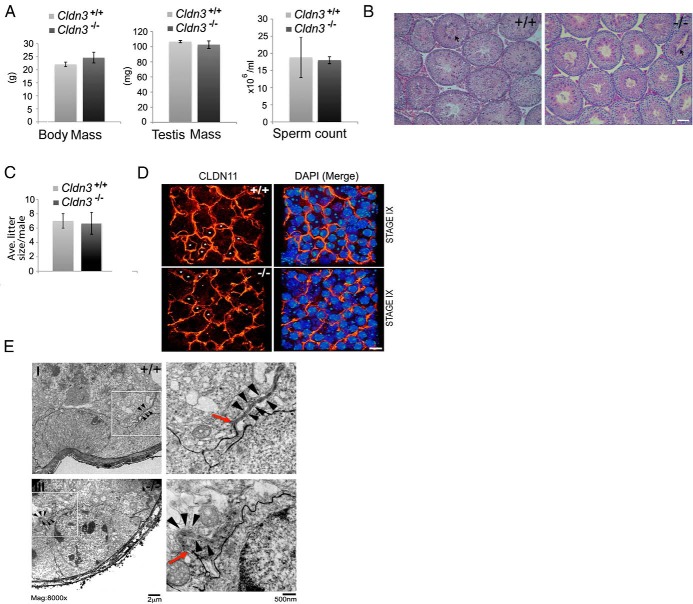
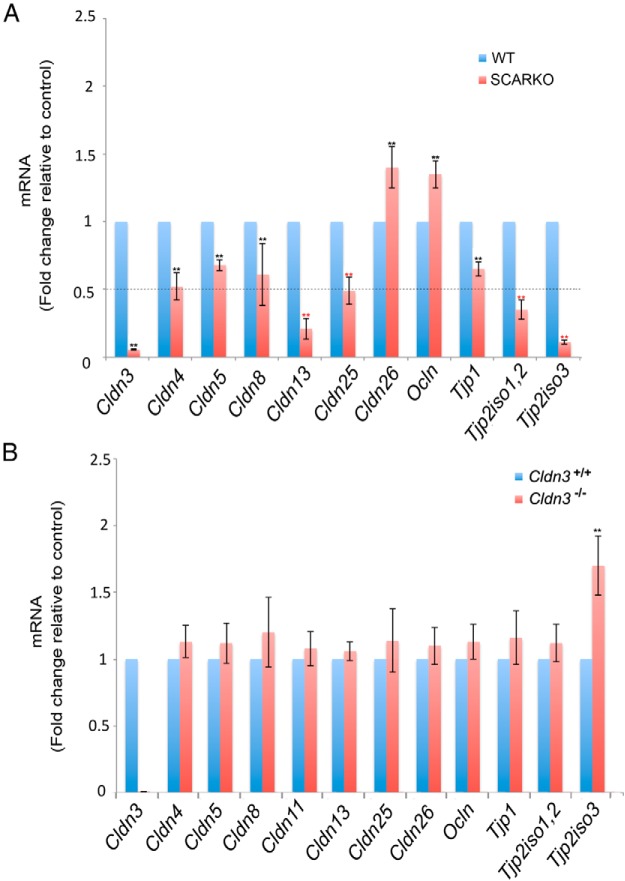
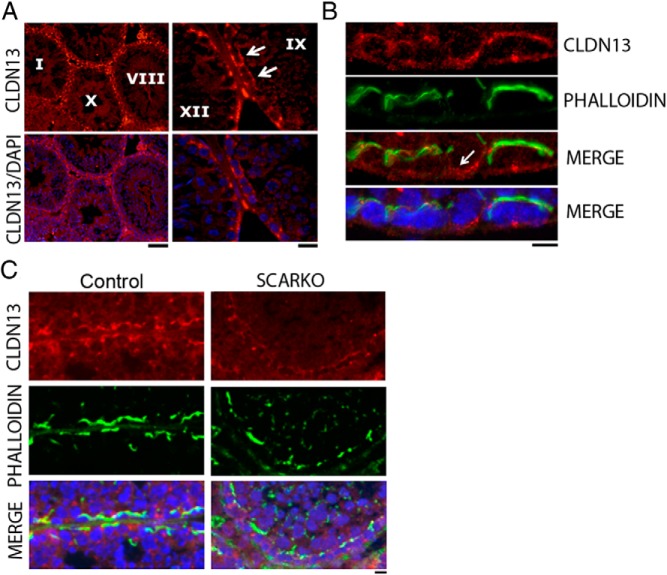
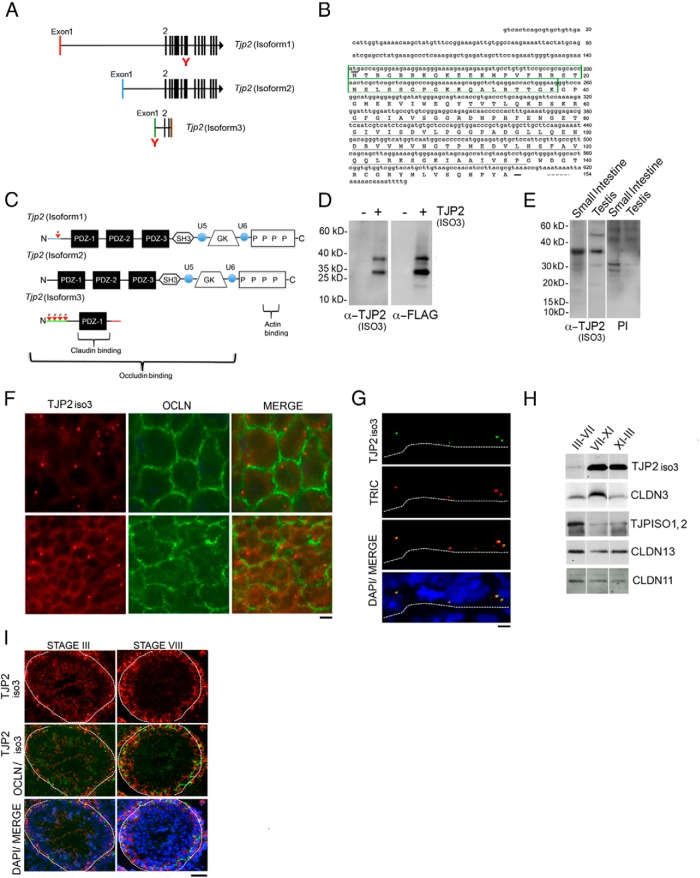
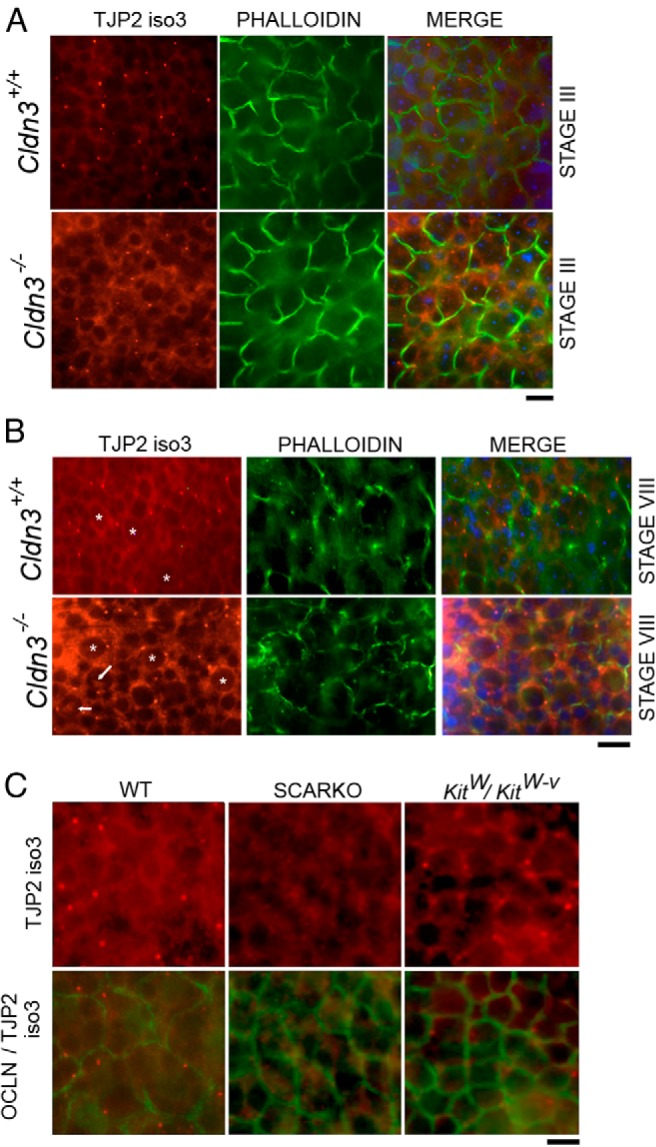
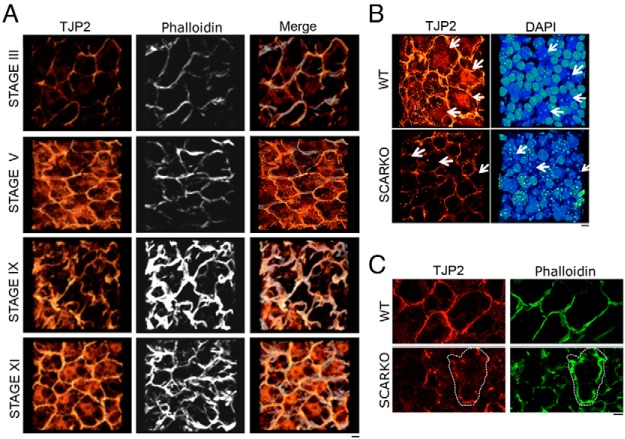
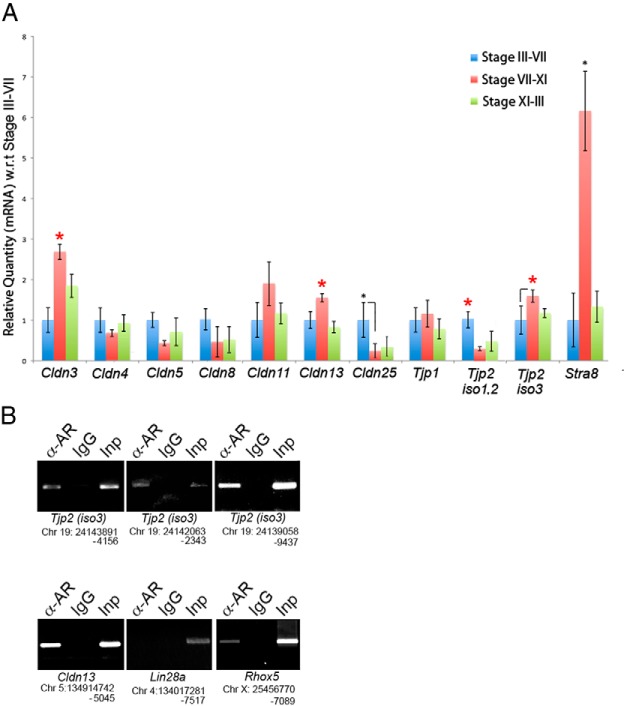
Sertoli cell tight junctions (SCTJs) of the seminiferous epithelium create a specialized microenvironment in the testis to aid differentiation of spermatocytes and spermatids from spermatogonial stem cells. SCTJs must be chronically broken and rebuilt with high fidelity to allow the transmigration of preleptotene spermatocytes from the basal to adluminal epithelial compartment. Impairment of androgen signaling in Sertoli cells perturbs SCTJ remodeling. Claudin (CLDN) 3, a tight junction component under androgen regulation, localizes to newly forming SCTJs and is absent in Sertoli cell androgen receptor knockout (SCARKO) mice. We show here that Cldn3-null mice do not phenocopy SCARKO mice: Cldn3(-/-) mice are fertile, show uninterrupted spermatogenesis, and exhibit fully functional SCTJs based on imaging and small molecule tracer analyses, suggesting that other androgen-regulated genes must contribute to the SCARKO phenotype. To further investigate the SCTJ phenotype observed in SCARKO mutants, we generated a new SCARKO model and extensively analyzed the expression of other tight junction components. In addition to Cldn3, we identified altered expression of several other SCTJ molecules, including down-regulation of Cldn13 and a noncanonical tight junction protein 2 isoform (Tjp2iso3). Chromatin immunoprecipitation was used to demonstrate direct androgen receptor binding to regions of these target genes. Furthermore, we demonstrated that CLDN13 is a constituent of SCTJs and that TJP2iso3 colocalizes with tricellulin, a constituent of tricellular junctions, underscoring the importance of androgen signaling in the regulation of both bicellular and tricellular Sertoli cell tight junctions.
Figures
References
-
- Dym M, Fawcett DW. The blood-testis barrier in the rat and the physiological compartmentation of the seminiferous epithelium. Biol Reprod. 1970;3:308–326. - PubMed
-
- Russell L. Movement of spermatocytes from the basal to the adluminal compartment of the rat testis. Am J Anat. 1977;148:313–328. - PubMed
-
- Mruk DD, Cheng CY. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis. Endocr Rev. 2004;25:747–806. - PubMed
-
- Tung KS, Teuscher C, Meng AL. Autoimmunity to spermatozoa and the testis. Immunol Rev. 1981;55:217–255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
