

SCF-FBXO31 E3 ligase targets DNA replication factor Cdt1 for proteolysis in the G2 phase of cell cycle to prevent re-replication
- PMID: 24828503
- PMCID: PMC4140253
- DOI: 10.1074/jbc.M114.559930
SCF-FBXO31 E3 ligase targets DNA replication factor Cdt1 for proteolysis in the G2 phase of cell cycle to prevent re-replication
Abstract
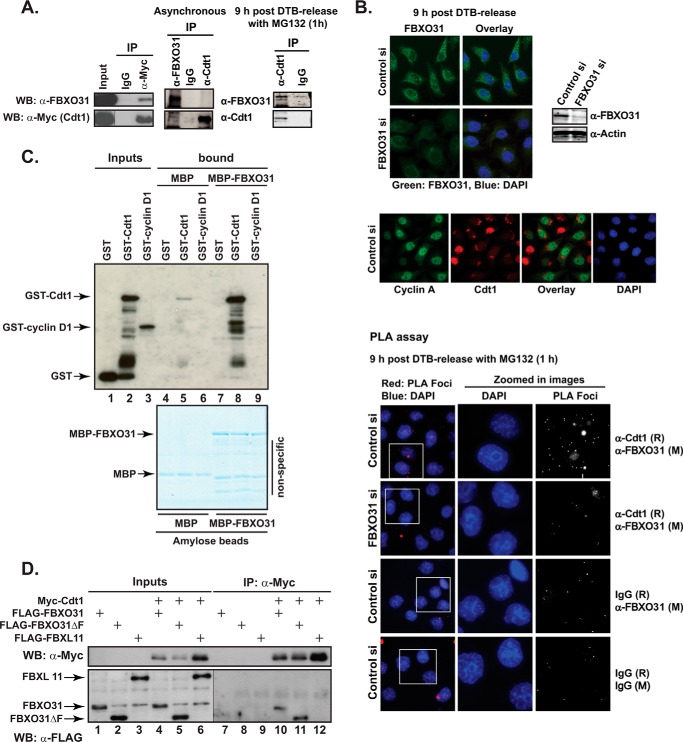
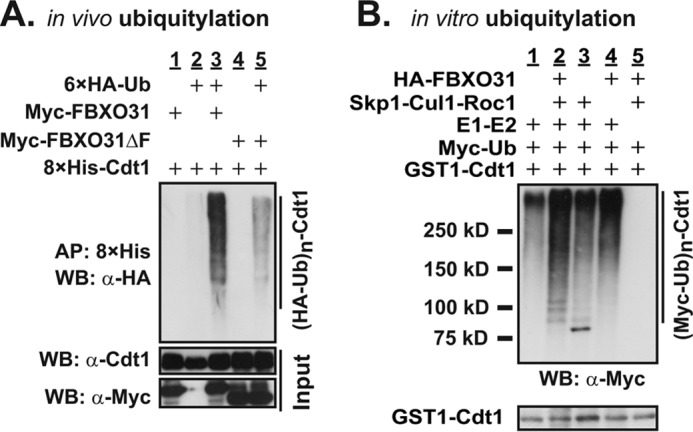
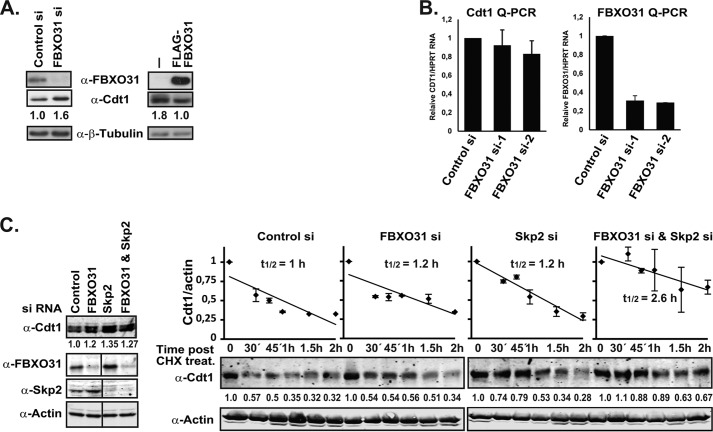
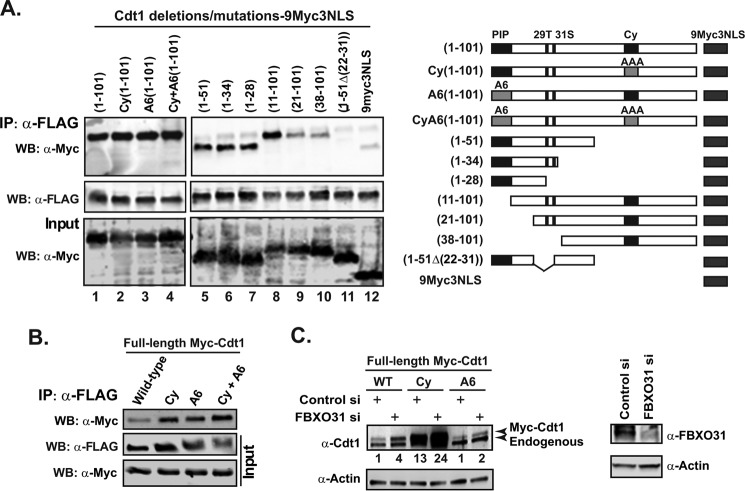
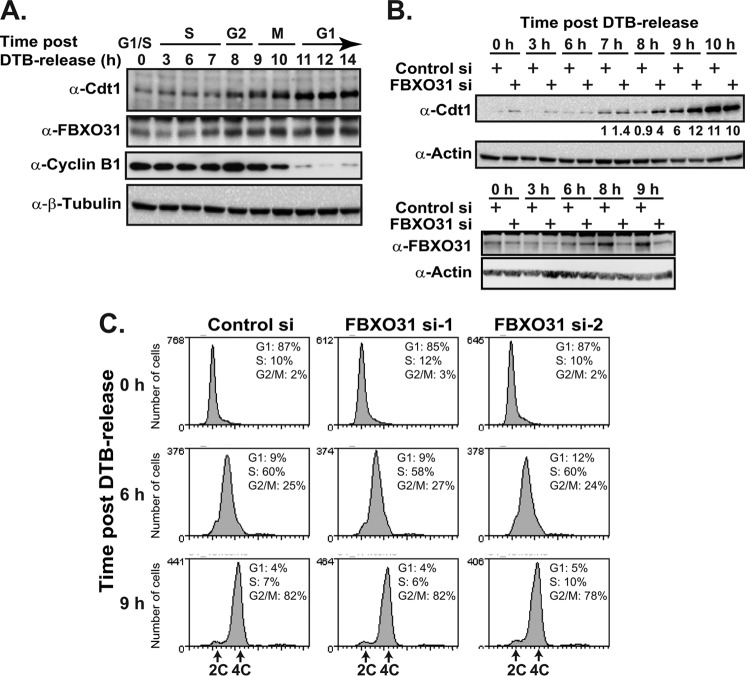
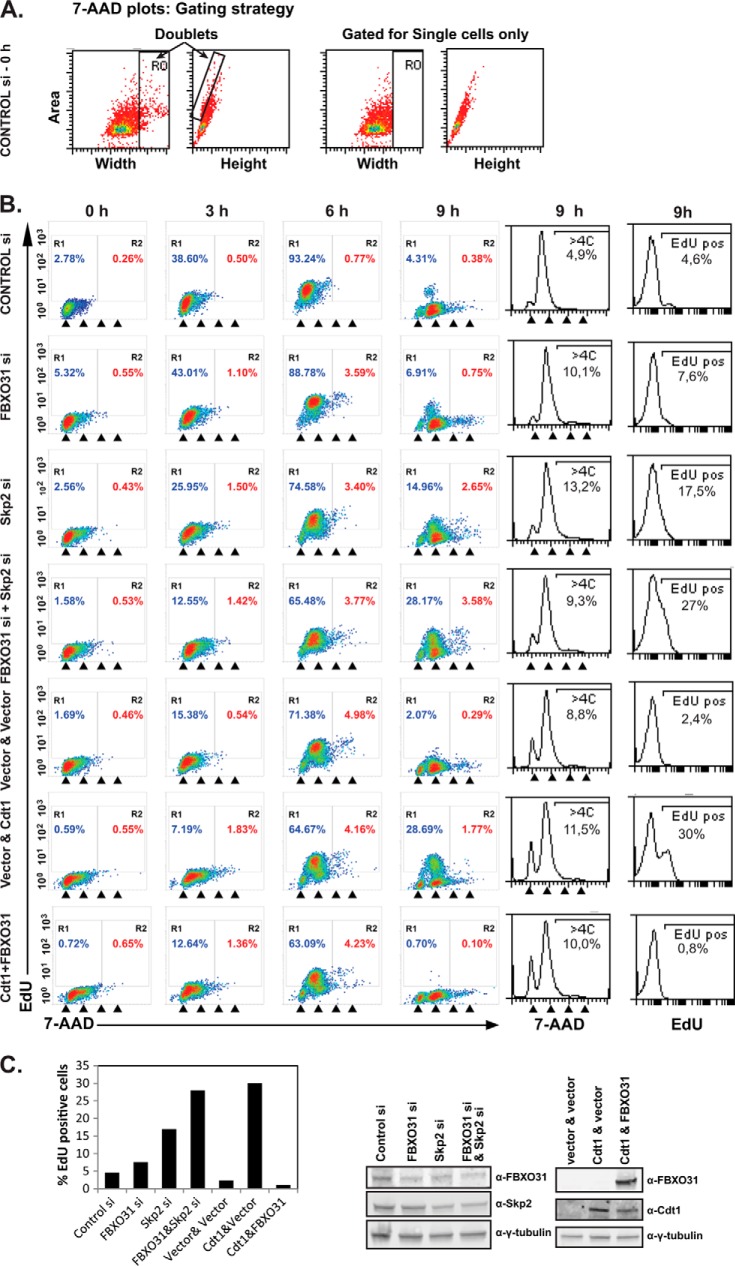
FBXO31 was originally identified as a putative tumor suppressor gene in breast, ovarian, hepatocellular, and prostate cancers. By screening a set of cell cycle-regulated proteins as potential FBXO31 interaction partners, we have now identified Cdt1 as a novel substrate. Cdt1 DNA replication licensing factor is part of the pre-replication complex and essential for the maintenance of genomic integrity. We show that FBXO31 specifically interacts with Cdt1 and regulates its abundance by ubiquitylation leading to subsequent degradation. We also show that Cdt1 regulation by FBXO31 is limited to the G2 phase of the cell cycle and is independent of the pathways previously described for Cdt1 proteolysis in S and G2 phase. FBXO31 targeting of Cdt1 is mediated through the N terminus of Cdt1, a region previously shown to be responsible for its cell cycle regulation. Finally, we show that Cdt1 stabilization due to FBXO31 depletion results in re-replication. Our data present an additional pathway that contributes to the FBXO31 function as a tumor suppressor.
Keywords: Cdt1; Cell Cycle; DNA Replication; E3 Ubiquitin Ligase; FBXO31; Genomic Instability; Protein Degradation.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
The tumor suppressor FBXO31 preserves genomic integrity by regulating DNA replication and segregation through precise control of cyclin A levels.J Biol Chem. 2019 Oct 11;294(41):14879-14895. doi: 10.1074/jbc.RA118.007055. Epub 2019 Aug 14. J Biol Chem. 2019. PMID: 31413110 Free PMC article.
-
The SCFFBXO46 ubiquitin ligase complex mediates degradation of the tumor suppressor FBXO31 and thereby prevents premature cellular senescence.J Biol Chem. 2018 Oct 19;293(42):16291-16306. doi: 10.1074/jbc.RA118.005354. Epub 2018 Aug 31. J Biol Chem. 2018. PMID: 30171069 Free PMC article.
-
FBXO31 protects against genomic instability by capping FOXM1 levels at the G2/M transition.Oncogene. 2017 Feb 16;36(7):1012-1022. doi: 10.1038/onc.2016.268. Epub 2016 Aug 29. Oncogene. 2017. PMID: 27568981
-
Regulation of DNA Replication Licensing and Re-Replication by Cdt1.Int J Mol Sci. 2021 May 14;22(10):5195. doi: 10.3390/ijms22105195. Int J Mol Sci. 2021. PMID: 34068957 Free PMC article. Review.
-
The Licensing Factor Cdt1 Links Cell Cycle Progression to the DNA Damage Response.Anticancer Res. 2020 May;40(5):2449-2456. doi: 10.21873/anticanres.14214. Anticancer Res. 2020. PMID: 32366388 Review.
Cited by
-
Control of DNA Replication Initiation by Ubiquitin.Cells. 2018 Sep 20;7(10):146. doi: 10.3390/cells7100146. Cells. 2018. PMID: 30241373 Free PMC article. Review.
-
The Toxoplasma gondii F-Box Protein L2 Functions as a Repressor of Stage Specific Gene Expression.PLoS Pathog. 2024 May 30;20(5):e1012269. doi: 10.1371/journal.ppat.1012269. eCollection 2024 May. PLoS Pathog. 2024. PMID: 38814984 Free PMC article.
-
THOC2 Mutations Implicate mRNA-Export Pathway in X-Linked Intellectual Disability.Am J Hum Genet. 2015 Aug 6;97(2):302-10. doi: 10.1016/j.ajhg.2015.05.021. Epub 2015 Jul 9. Am J Hum Genet. 2015. PMID: 26166480 Free PMC article.
-
Ubiquitin-specific protease 53 promotes osteogenic differentiation of human bone marrow-derived mesenchymal stem cells.Cell Death Dis. 2021 Mar 4;12(3):238. doi: 10.1038/s41419-021-03517-x. Cell Death Dis. 2021. PMID: 33664230 Free PMC article.
-
Deregulation of F-box proteins and its consequence on cancer development, progression and metastasis.Semin Cancer Biol. 2016 Feb;36:33-51. doi: 10.1016/j.semcancer.2015.09.015. Epub 2015 Sep 30. Semin Cancer Biol. 2016. PMID: 26432751 Free PMC article. Review.
References
-
- Härkönen P., Kyllönen A. P., Nordling S., Vihko P. (2005) Loss of heterozygosity in chromosomal region 16q24.3 associated with progression of prostate cancer. Prostate 62, 267–274 - PubMed
-
- Kumar R., Neilsen P. M., Crawford J., McKirdy R., Lee J., Powell J. A., Saif Z., Martin J. M., Lombaerts M., Cornelisse C. J., Cleton-Jansen A. M., Callen D. F. (2005) FBXO31 is the chromosome 16q24.3 senescence gene, a candidate breast tumor suppressor, and a component of an SCF complex. Cancer Res. 65, 11304–11313 - PubMed
-
- Launonen V., Mannermaa A., Stenbäck F., Kosma V. M., Puistola U., Huusko P., Anttila M., Bloigu R., Saarikoski S., Kauppila A., Winqvist R. (2000) Loss of heterozygosity at chromosomes 3, 6, 8, 11, 16, and 17 in ovarian cancer: correlation to clinicopathological variables. Cancer Genet. Cytogenet. 122, 49–54 - PubMed
-
- Lin Y. W., Lee I. N., Chen C. H., Huang G. T., Lee H. S., Lee P. H., Lu F. J., Sheu J. C. (2001) Deletion mapping of chromosome 16q24 in hepatocellular carcinoma in Taiwan and mutational analysis of the 17-β-HSD gene localized to the region. Int. J. Cancer 93, 74–79 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
