

Retinoic acid-related orphan receptor γ (RORγ): a novel participant in the diurnal regulation of hepatic gluconeogenesis and insulin sensitivity
- PMID: 24831725
- PMCID: PMC4022472
- DOI: 10.1371/journal.pgen.1004331
Retinoic acid-related orphan receptor γ (RORγ): a novel participant in the diurnal regulation of hepatic gluconeogenesis and insulin sensitivity
Abstract
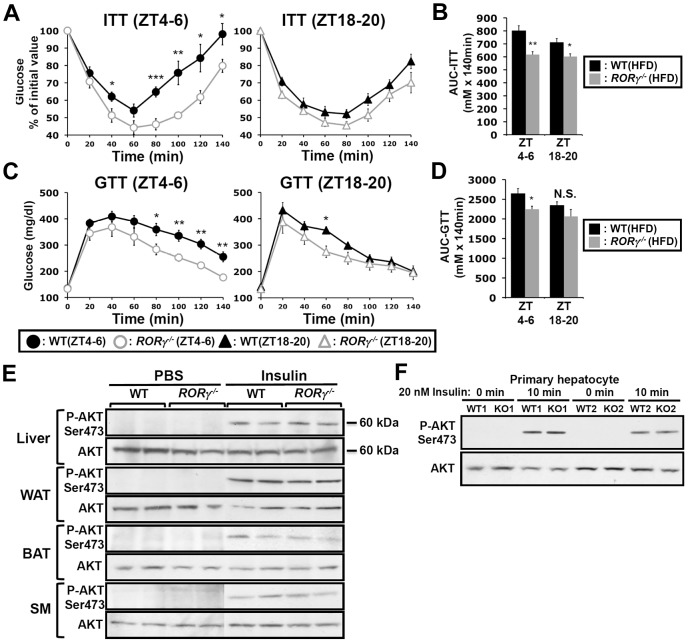
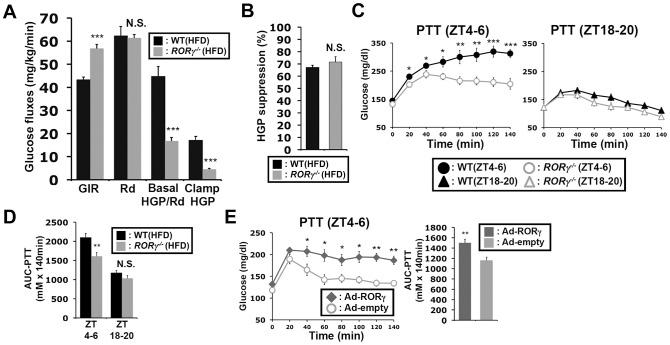
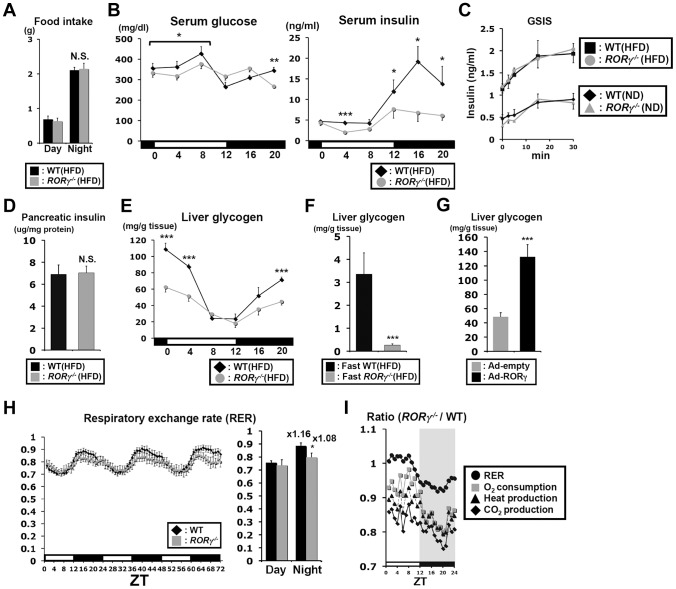
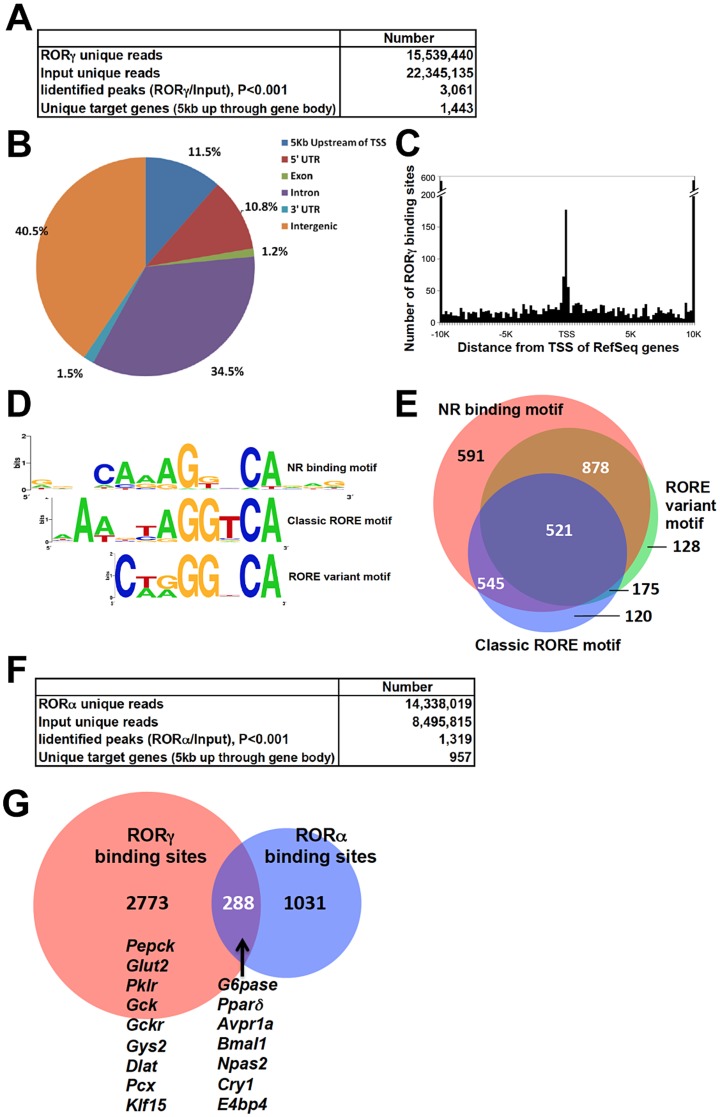
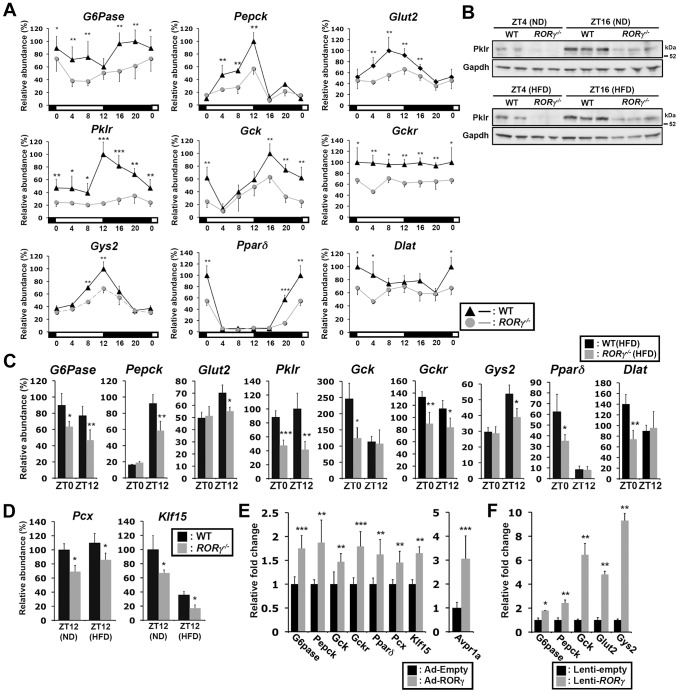
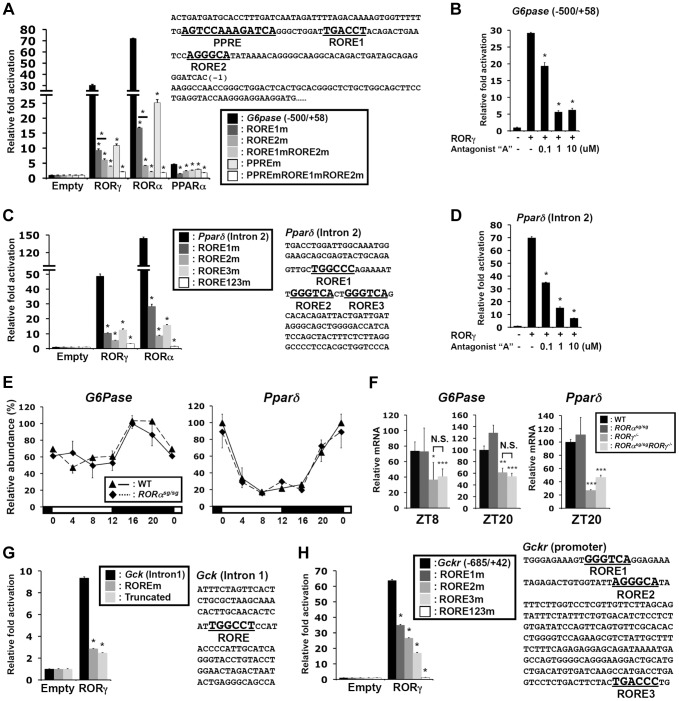
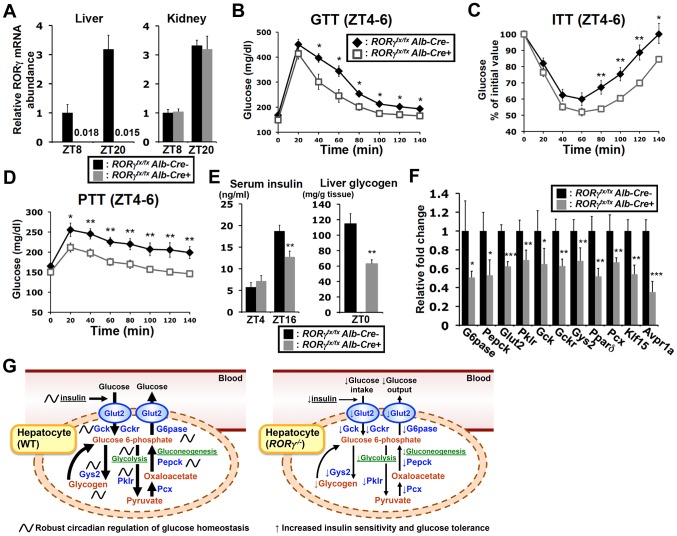
The hepatic circadian clock plays a key role in the daily regulation of glucose metabolism, but the precise molecular mechanisms that coordinate these two biological processes are not fully understood. In this study, we identify a novel connection between the regulation of RORγ by the clock machinery and the diurnal regulation of glucose metabolic networks. We demonstrate that particularly at daytime, mice deficient in RORγ exhibit improved insulin sensitivity and glucose tolerance due to reduced hepatic gluconeogenesis. This is associated with a reduced peak expression of several glucose metabolic genes critical in the control of gluconeogenesis and glycolysis. Genome-wide cistromic profiling, promoter and mutation analysis support the concept that RORγ regulates the transcription of several glucose metabolic genes directly by binding ROREs in their promoter regulatory region. Similar observations were made in liver-specific RORγ-deficient mice suggesting that the changes in glucose homeostasis were directly related to the loss of hepatic RORγ expression. Altogether, our study shows that RORγ regulates several glucose metabolic genes downstream of the hepatic clock and identifies a novel metabolic function for RORγ in the diurnal regulation of hepatic gluconeogenesis and insulin sensitivity. The inhibition of the activation of several metabolic gene promoters by an RORγ antagonist suggests that antagonists may provide a novel strategy in the management of metabolic diseases, including type 2 diabetes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Fitzsimmons RL, Lau P, Muscat GE (2012) Retinoid-related orphan receptor alpha and the regulation of lipid homeostasis. J Steroid Biochem Mol Biol 130: 159–168. - PubMed
-
- Eberl G, Littman DR (2003) The role of the nuclear hormone receptor RORgammat in the development of lymph nodes and Peyer's patches. Immunol Rev 195: 81–90. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
